Математика Теормех Сопромат Образцы решений
МАДИ (дневное и вечернее отделения)
Математика
РГР 1.1. Начала математичесткого анализа (здесь примеры по старой методичке, до 2015 года).
Номер 2.5
Номер 10.10
Номер 20.22
РГР 1.2. Линейная алгебра и аналитическая геометрия.
Номер 10.3
Номер 14.13
Номер 18.28
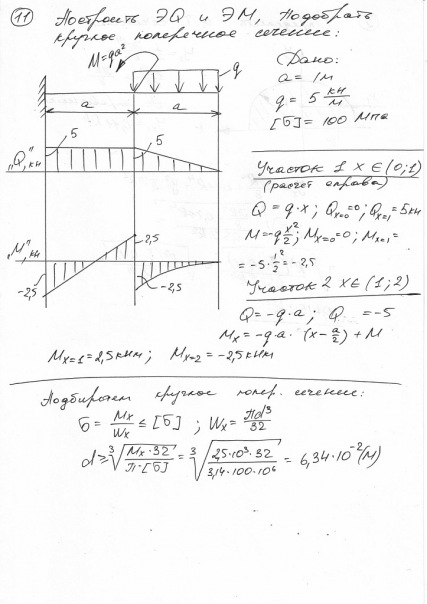
Номер 25.16
РГР 2.1. Неопределенные интегралы.
Номер 5.1
Номер 13.15
Номер 20.26
Номер 27.22
РГР 2.
Номер 3.9
Номер 8.17
Номер 13.26
Номер 20.30
РГР 3.1. Дифференциальные уравнения.
Номер 4.29
Номер 6.20
Номер 14.12
Номер 18.2
РГР 3.2. Ряды.
Номер 5.7
Номер 10.26
Номер 16.28
Номер 20.15
РГР Теория вероятностей и математическая статистика (бывшая 4.1).
Номер 9.6
Номер 15.1
-
Номер 19.11
Номер 26.21
Номер 33.
 30
30
Теоретическая механика
Задача К2 (Вариант 4 пакет 3, соответствует схеме 28 варианта по методичке Ермакова)
Задача К3.1 (Вариант 7 пакет 2, соответствует схеме 22 варианта по методичке Ермакова)
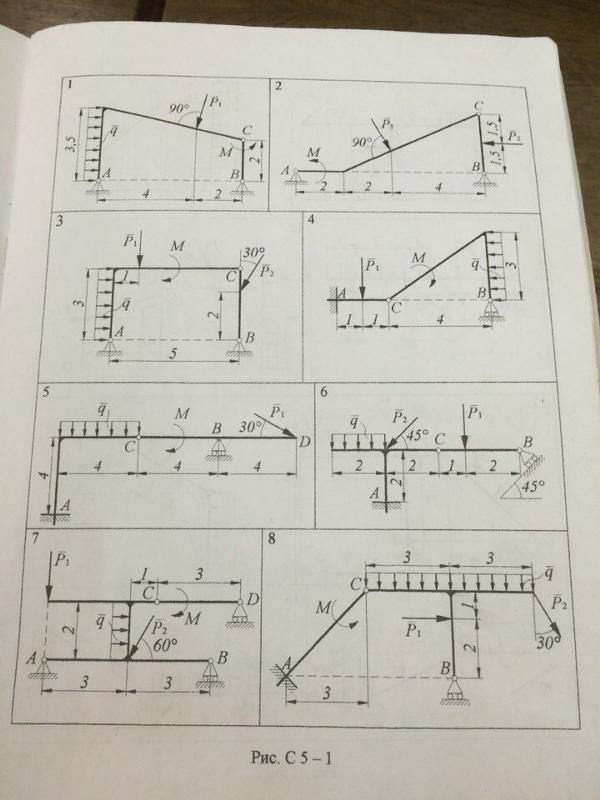
Задача С3 (Вариант 16 по методичке Ермакова)
Задача С5 (Вариант 23 по методичке Ермакова)
Задача Д4 (Вариант 4 по методичке Ермакова)
Задача Д5.2 (Вариант 10 по методичке Ермакова)
Задача Д10 (Вариант 25 по методичке Ермакова)
Сопромат
Задание 1. Определение внутренних сил. Расчет на действие продольных сил.
Определение внутренних сил. Расчет на действие продольных сил.
Задача 1.2 Вариант подчеркнут
Задача 1.3 Вариант подчеркнут
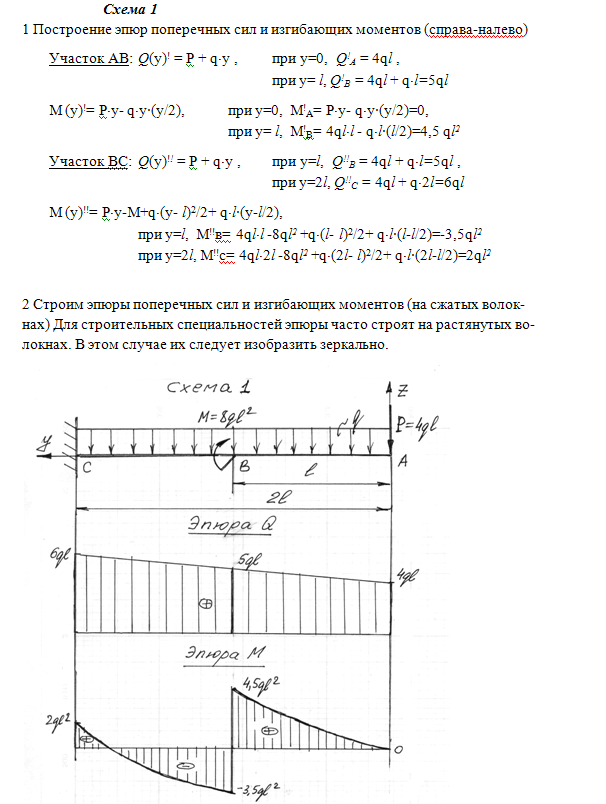
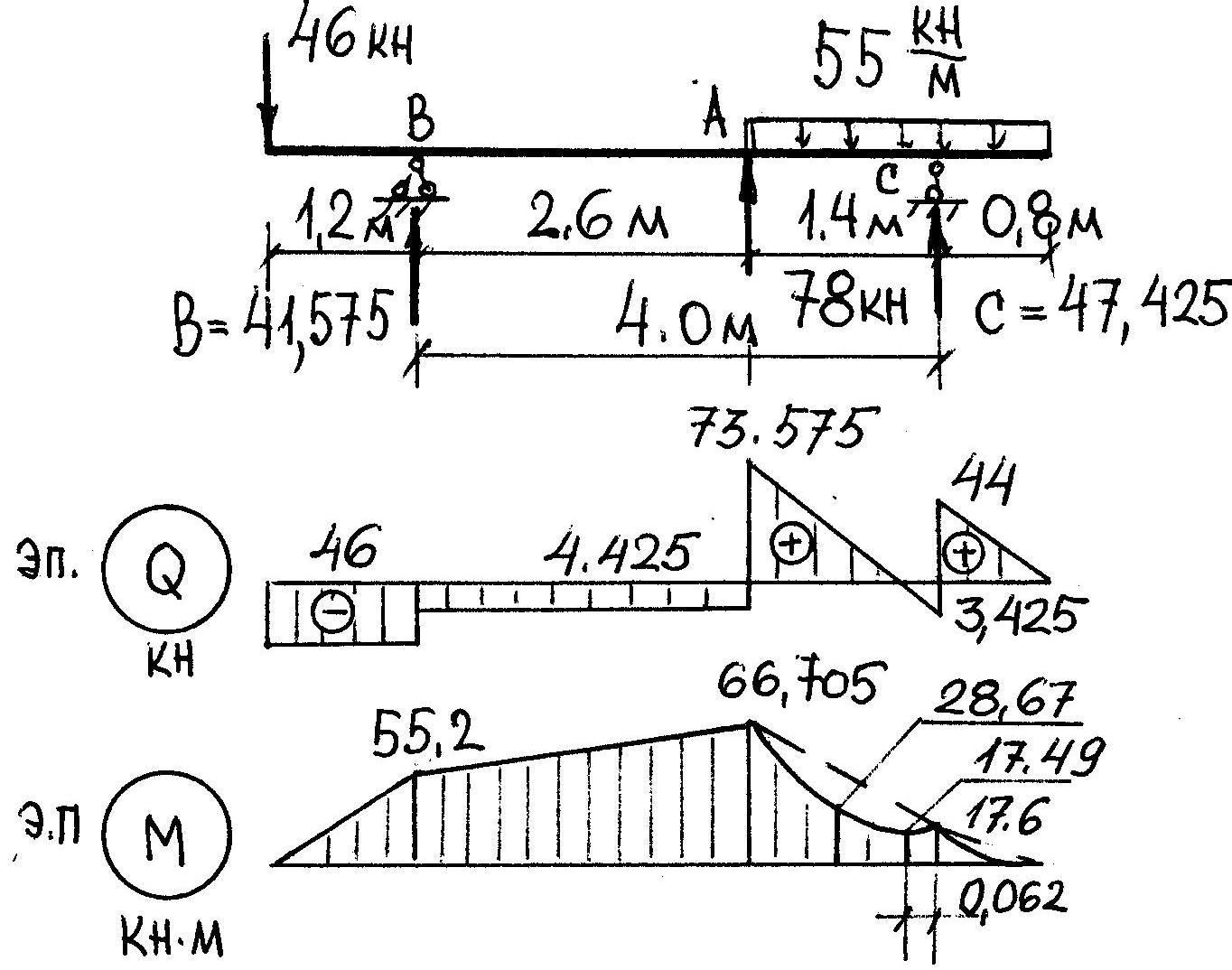
Задание 3. Расчет балки на изгиб.
Задание 3. Схемы 2 и 3. Вариант подчеркнут
Второй семестр изучения сопромата. Задания со стенда. Задача 2.3. Неразрезная балка. Вариант 26
Строительная механика
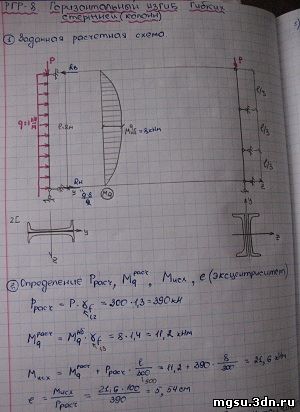
Задание 1. Задача 2. Расчет статически определимой фермы. Вариант 8. Данные отмечены
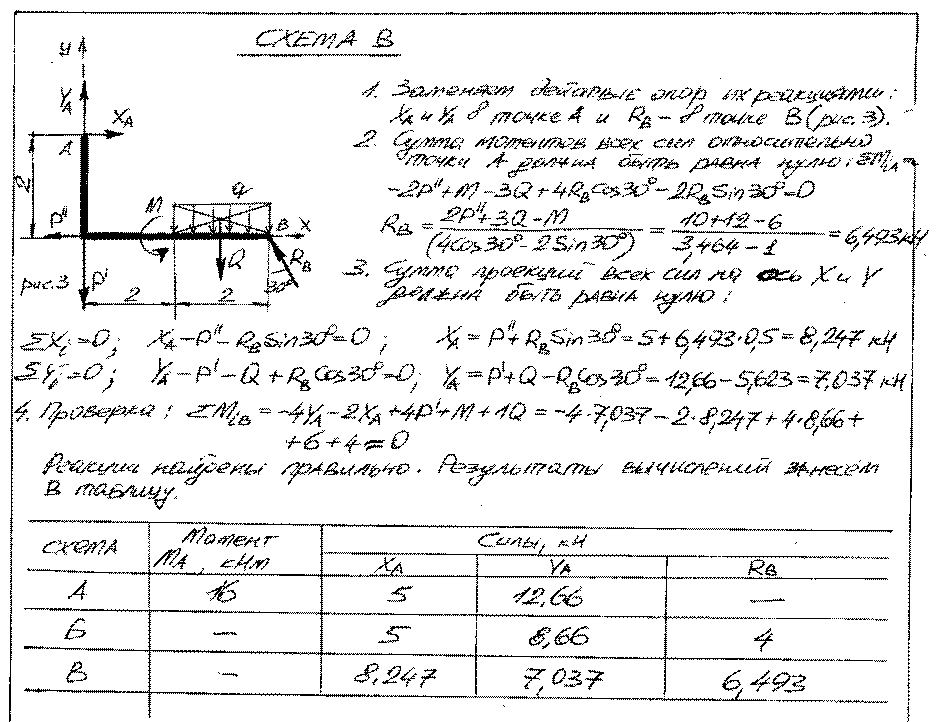
Задание 6. Расчет плоских статически неопределимых систем на подвижную нагрузку
Задание 6. Решение. Вариант подчеркнут
МАДИ (Заочное отделение)
В настоящее время раздел находится в разработке.
Математика
Методичка Мироненко. К/Р № 2. Вариант №7
Методичка Мироненко. К/Р № 5. Вариант №2
Теоретическая механика
Методичка Ермакова (№ 677) для дневного отделенияМетодичка для студентов заочного отделения
Задача С3. Вариант 13. (По методичке для студентов заочного отделения)
Задача К2. Вариант 28. (По методичке для студентов заочного отделения)
Задача Д1. Вариант 8. (По методичке для студентов заочного отделения)
Сопромат
Первый семестр сопромата. Задача 3.1. Вариант 12. Группа 2ЗАТ2. Год поступления 2012.
Строительная механика
Первый семестр строительной механики.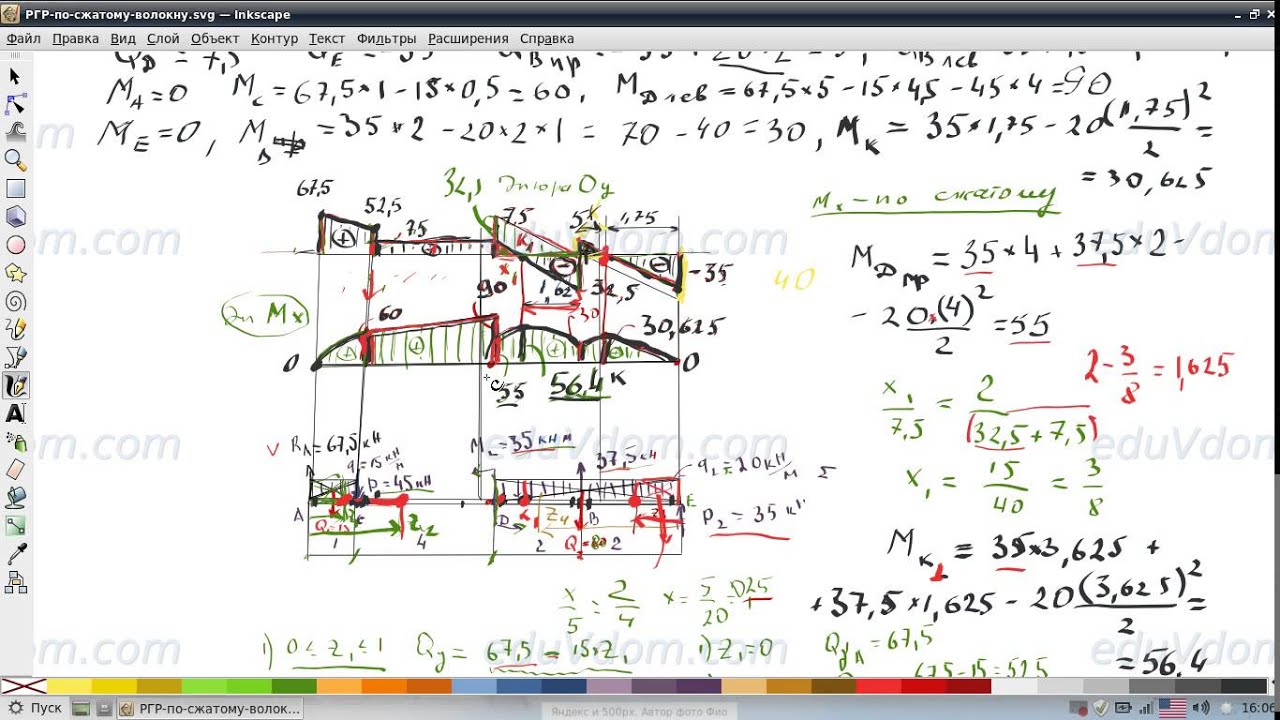 Задача 1. Вариант 9.
Задача 1. Вариант 9.
РГР на заказ по термеху, сопромату, электротехнике
Расчетно-графическая работа – это весьма специфический вид задания, который характерен для студентов технических направлений обучения. Наиболее частыми являются ргр по сопромату (сопротивление материалов), термеху (технической механике), электротехнике, прикладной механике, ТОЭ (теоретические основы электротехники) и т.д.
Сама суть работы включает в себя сложные расчеты и отображение результатов этих расчетов в графическом виде с точными и подробными объяснениями всех деталей и нюансов работы.Сколько стоит написание РГР?
Стоимость РГР в Оренбурге — от 99 ₽. Сроки выполнения — от 1 дня.
Бесплатные доработки. Прохождение проверки на уникальность. Гарантия выполнения или возврат денег.
Без определенных знаний и навыков выполнения подобных заданий, впервые столкнувшись с необходимостью самостоятельно подготовить материал, студент может растеряться.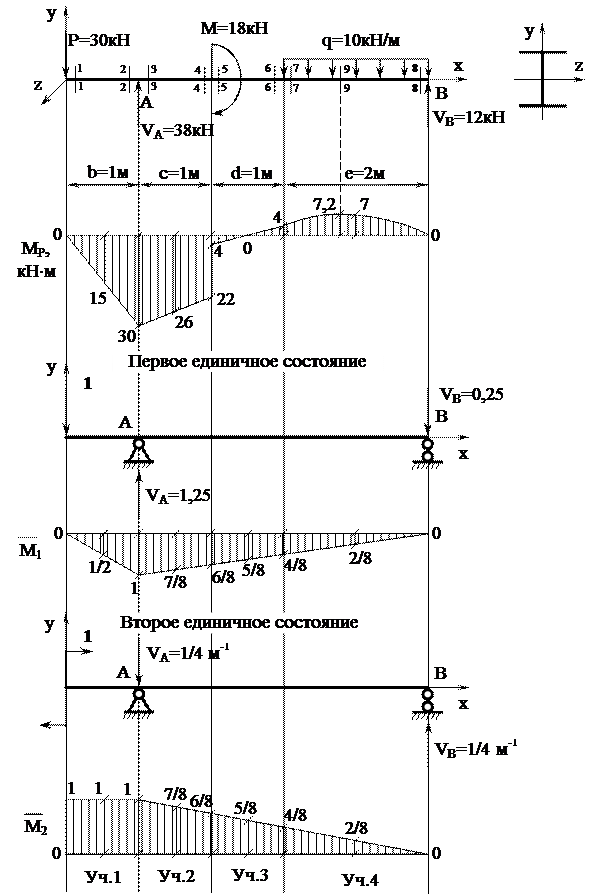 Так как же поступить, к кому обратиться за помощью? Есть несколько вариантов развития событий. Первый- самостоятельно выполнить все так, как посчитает нужным. Однако в этом случае, полной уверенности в том, что все сделано правильно, быть не может. Второе, не менее опасное решение — можно пойти к студентам-старшекурсникам и взять готовый материал у них. Однако здесь также никаких гарантий того, что работа будет выполнена правильно, а все расчеты произведены верно.
Так как же поступить, к кому обратиться за помощью? Есть несколько вариантов развития событий. Первый- самостоятельно выполнить все так, как посчитает нужным. Однако в этом случае, полной уверенности в том, что все сделано правильно, быть не может. Второе, не менее опасное решение — можно пойти к студентам-старшекурсникам и взять готовый материал у них. Однако здесь также никаких гарантий того, что работа будет выполнена правильно, а все расчеты произведены верно.
Третий и самый обдуманный и взвешенный вариант – это обратиться за помощью к специалистам, которые не только имеют богатый опыт выполнения подобных заявок, но и достаточно глубокие знания для того, чтобы сделать работу профессионально, а расчеты произвести максимально точно.
Выполнить расчетно-графическую работу срочно и недорого
РГР на заказ у специалистов – это материал, который будет выполнен с учетом всех требований и методических рекомендаций. Каждое графическое изображение будет подкреплено уточняющими сопроводительными записками, содержащими формулы и все необходимые расчеты.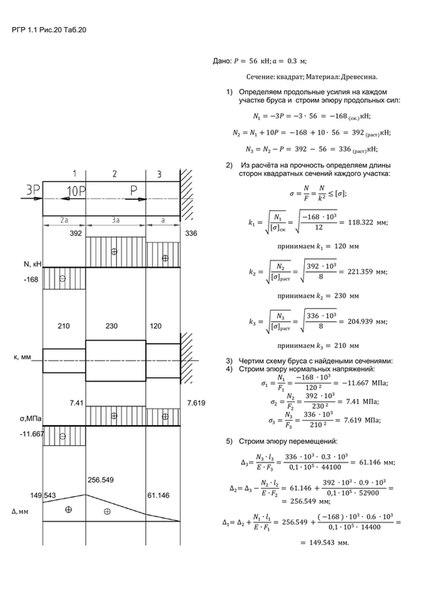 Мы готовы принять к выполнению как простую ргр, содержащую 1-2 графические изображения, так и объемные задания, отнимающие много времени и содержащие сложные расчеты.
Мы готовы принять к выполнению как простую ргр, содержащую 1-2 графические изображения, так и объемные задания, отнимающие много времени и содержащие сложные расчеты.
Стоимость расчетно-графической работы на заказ вполне доступна для простого студента и напрямую зависит от сложности задания и его объема.
Мы принимаем к выполнению «срочные» заявки, однако стоит понимать, что данное задание не может быть выполнено за несколько часов, да и за срочность необходимо будет заплатить. Поэтому рекомендуем Вам при малейших сомнениях в том, что Вы самостоятельно одолеете данный труд, сразу же обращаться за помощью к нашим сотрудникам, и тогда успех Вам гарантирован.
Учебно-методические материалы Кафедры механики деформируемого твердого тела
В этом разделе Вы можете скачать электронные версии учебных пособий, методических указаний и бланки заданий на выполнение РГР, созданные преподавателями кафедры. Также есть возможность воспользоваться учебными пособиями, созданными в других вузах России и рекомендованными кафедрой для использования в учебном процессе, и справочным материалом, используемым при изучении курсов кафедры.
Учебные пособия и методические работы для студентов, обучающихся по программе магистра
- Иванников Л.М. Решение двумерной задачи теплопроводности методом конечных элементов в MATHCAD. Методические указания при изучении курса «Аналитические и численные методы решения уравнений математической физики» скачать (747.1 KБ)
- Дойхен Ю.М. Ким Т.С. Тен Ен Со Расчет рам на устойчивость и динамические нагрузки. Методические указания при изучении курса «Динамика и устойчивость зданий и сооружений» скачать (21.7 MБ)
- Тен Ен Со Решение задач теории упругости с применением Mathcad 14.0. Учебное пособие для студентов обучающихся в направлении строительных специальностей, при изучении курса «теории упругости» скачать (932.2 KБ)
- Тен Ен Со Программы Mathcad 14.0 к учебному пособию по «теории упругости» скачать (1.7 MБ)
- Лукашевич А.А. Современные численные методы строительной механики. Учебное пособие скачать (1.8 MБ)
- Иванников Л.
 М. Расчет пространственных ферм методом конечных элементов в Mathcad. Методические указания и контрольные задания к выполнению лабораторной работы по курсу «Аналитические и численные методы решения уравнений математической физики» скачать (869.3 KБ)
М. Расчет пространственных ферм методом конечных элементов в Mathcad. Методические указания и контрольные задания к выполнению лабораторной работы по курсу «Аналитические и численные методы решения уравнений математической физики» скачать (869.3 KБ) - В.В. Иовенко, Л.М. Иванников. Расчет составных сосудов. Методические указания по курсу «Сопротивление материалов» для студентов механических и строительных специальностей дневной формы обучения, для учебно-исследовательской работы, для магистров скачать (842.5 KБ)
Учебные пособия и другие методические работы для студентов дневной формы обучения
- Потапова Л.Б. Сложное сопротивление. Статический и динамический расчет вала. Методические указания к выполнению расчетно-графической работы по курсу «Сопротивление материалов» для механических специальностей скачать (821.4 KБ)
- Потапова Л.Б. Сопротивление материалов: научно-исследовательская работа скачать (700.6 KБ)
- Потапова Л.
 Б., Ярцев В.П. Механика материалов при сложном напряженном состоянии скачать (2.6 MБ)
Б., Ярцев В.П. Механика материалов при сложном напряженном состоянии скачать (2.6 MБ) - Вайсфельд А.А. Основы строительной механики. Учебное пособие для студентов специальностей «Архитектура» и «Дизайн архитектурной среды» скачать (2.0 MБ)
- А. Д. Ловцов Метод конечных элементов. Методические указания и варианты заданий к выполнению лабораторных работ №1 и №2 для студентов строительных специальностей скачать (564.0 KБ)
- А. Д. Ловцов Метод конечных элементов. Методические указания и варианты заданий к выполнению лабораторных работ №3 и №4 для студентов строительных специальностей скачать (652.1 KБ)
В.Е.Киселев Метод конечных элементов. Методические указания и варианты заданий к выполнению лабораторных работ №5 и №6 для студентов строительных специальностей скачать (615.6 КБ)
- А. А. Лукашевич, А. А. Вайсфельд. Варианты заданий к выполнению расчетно-графических работ по курсу строительной механики для специальностей «Архитектура» и «Дизайн архитектурной среды» скачать (3.
 2 MБ)
2 MБ)
Учебные пособия и методические работы для студентов ускоренной и заочной форм обучения
- Шестаков И.А. Сопротивление материалов. Часть II. Внецентренное сжатие. Изгиб с кручением. Устойчивость сжатого стержня. Методические указания к выполнению контрольных работ скачать (572.1 KБ)
- Киселев В.Е. Строительная механика. Часть 1. Методические указания к выполнению контрольных работ по курсу «Строительная механика» скачать (630.3 KБ)
- Дойхен Ю.М., Лукашевич А.А., Потапова Л.Б. Сопротивление материалов. Методические указания и контрольные задания скачать (438.9 KБ)
- А.А. Вайсфельд, В.Е. Киселев, А.Д. Ловцов. Строительная механика. Часть II. Примеры выполнения контрольных работ скачать (1.6 MБ)
- В. Е. Киселев Анализ неизменяемости сооружений. Методические указания к изучению раздела курса «Строительная механика» для студентов строительных специальностей скачать (367.7 KБ)
- А.А. Вайсфельд, Л.М. Иванников, В.
 Е. Киселев, А.Д. Ловцов, А.А. Лукашевич. Строительная механика. Часть I. Методические указания и контрольные задания для студентов строительных специальностей скачать (886.4 KБ)
Е. Киселев, А.Д. Ловцов, А.А. Лукашевич. Строительная механика. Часть I. Методические указания и контрольные задания для студентов строительных специальностей скачать (886.4 KБ) - В.В. Иовенко. Примеры решения задач контрольных работ по сопротивлению материалов. Методические указания по курсу «Сопротивление материалов» скачать (764.0 KБ)
- В.В. Иовенко. Основные сведения по курсу «Сопротивление материалов» скачать (148.2 KБ)
- В.В. Иовенко. Краткий курс лекций по курсу «Сопротивление материалов» для студентов сокращенной формы обучения специальностей ХТПЭ, ХПД, ООС и заочной и дистанционной форм обучения скачать (2.7 MБ)
Учебные пособия, рекомендованные кафедрой к использованию в учебном процессе
- Windjview программа для просмотра файлов формата djvu скачать (605.1 KБ)
- Дарков А.В. Шапошников Н.Н. Строительная механика скачать (6.9 MБ)
- Шапин В.И. Прикладная механика скачать (688.1 KБ)
- Шапин В.
 И. Сопротивление материалов скачать (1.4 MБ)
И. Сопротивление материалов скачать (1.4 MБ) - Чупин В.В., Черногубов Д.Е. Расчет на устойчивость сжатых стержней скачать (243.0 KБ)
- Мущанов В.Ф., Жук Н.Р. Строительная механика. Конспект лекций скачать (178.8 KБ)
- Коперник Г.Р., Козлов В.В. Определение перемещений в статически определимых системах скачать (395.7 KБ)
- Ю. Г. Плотников. Матрицы в строительной механике. Учебное пособие скачать (2.3 MБ)
- А.С.Волков, Ю.Г. Плотников Динамические расчеты упругих систем. Учебное пособие скачать (2.0 MБ)
Справочные материалы
Учебно-методические указания и пособия по Теоретической механике
- Задохин Б.С. Теоретическая механика. Учебное пособие. Хабаровск, ХГТУ, 1998.- 72 c. 2. Лукашевич Н.К. Теоретическая механика: тексты лекций. Хабаровск: Изд-во Тихоокеан. гос. ун-та, 2009.- 263 с.
- Теоретическая механика: методические указания и контрольные задания для студентов-заочников/ Под ред.
 С.М. Тарга М.: Высшая школа, 1989. 111 с.
С.М. Тарга М.: Высшая школа, 1989. 111 с.
Список основной литературы по Теоретической механике
- Бутенин Н.В., Лунц Я.Л., Меркин Д.Р. Курс теоретической механики. ч.1., М.: Наука, 1985. 240 с.
- Бутенин Н.В., Лунц Я.Л., Меркин Д.Р. Курс теоретической механики. ч.2., М.: Наука, 1985. 496 с.
- Яблонский А.А., Никифорова В.М. Курс теоретической механики: Учеб. пособие для студ. вузов (гриф МО): Статика. Кинематика. Динамика – 8-е изд. стереотип. – СПб: Лань, 2001. – 768 с.
- Павловский М.А., Путята Т.В. Теоретическая механика. Киев.: Вища школа, 1985, 329 с.
- Тарг С.М. Краткий курс теоретической механики: Учебник для студ. втузов (гриф МО). – 12-е изд., стереотип. – М.: Высш. шк., 2002. – 416 с.
Входной контроль знаний
Входной контроль знаний студентов включает в себя задачи:
- Расчет векторного треугольника с применением теорем синуса и косинуса
- Определение проекций вектора на ось плоской и пространственной систем координат
- Вычисление скалярного произведения векторов, по их известным координатам в декартовой прямоугольной координатной системе
- Вычисление векторного произведения векторов
- Расчет векторного уравнения с целью определения величин неизвестных векторов
Образец входного контроля по теоретической механике скачать (135.
 3 KБ)
3 KБ)Упражнения
Разложение вектора силы на составляющие его проекции Fx и Fy
Упражнение «Определение проекции силы на координатную ось» скачать (88.5 KБ)
Вычисления алгебраического значения момента силы относительно точки.
Упражнение «Определение момента силы относительно точки» скачать (148.5 KБ)
Знание связей и их реакций.
Таблица «Связи и их реакции» скачать (157.4 KБ)
Составление уравнений равновесия для плоской произвольной системы сил.
Определение реакций связей твердого тела скачать (172.5 KБ)
Общее уравнение динамики
Лекция скачать (203.0 KБ)
Лекции по Теоретической механике
Лекции по дисциплине скачать (724.2 KБ)
Экзаменационные вопросы
Для cпециальности УИ-01(у) скачать (117.4 KБ)
Для групп С-11, С-12, С-13, НД-11, СУЗ-11 скачать (116.0 KБ)
Пример экзаменационных задач
Для cпециальностей ПГС, ГСХ, ТВ, ВВ, ПГС(у) скачать (171.
 0 KБ)
0 KБ)Для cпециальностей ААХ, СЭМ, ДВС, ЭСУ скачать (238.1 KБ)
| http://alhimikov.net | Полезное по химии. | Химия: решение задач | Решение задач и контрольных работ по химии, химический софт, лекции и учебники в электронном формате. |
Помощь по химии | Решаем задачи по химии, а также отвечаем на любые другие вопросы химической тематики |
Поможем решить задачи, ИДЗ, РГР, контрольные по следующим предметам: Математика, Физика, Теоретическая механика, Химия, Строймех, ТОЭ, Сопромат и.т.д. все задачи. |
http://sm.teormex.net/ |
Решение задач по сопромату, шпоры |
Термех Тарга С.М | Всех разделов, всех годов.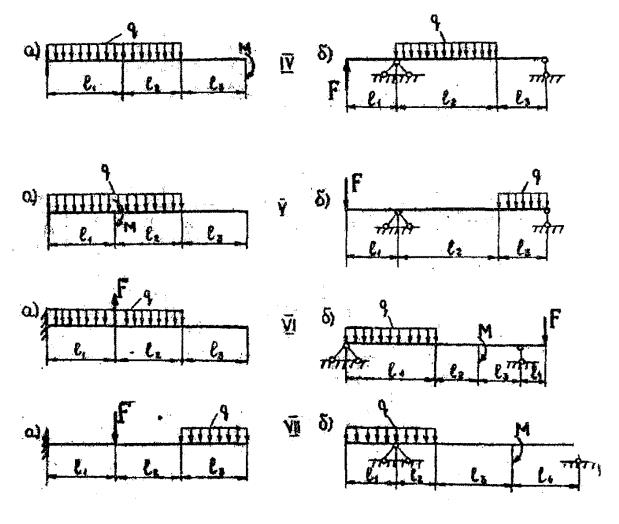 Решение задач других авторов, самые низкие цены. Решение задач других авторов, самые низкие цены.
|
Новости образования, база данных учебных заведений, справочники школ, вузов, техникумов, колледжей, советы для поступающих в вузы, тесты ЕГЭ. | Термех.NET |
Поверенные решения задач теоретической механики (термеха) из задачника Яблонского А.А. (1985 г.) и Мещерского И.В (1986 г.). |
Математика для школы. Понятные объяснения .Решение задач и примеров по математике. | |
Официальный сайт школы №89 г. Казани. Информация о школе, учениках, учителях, страница для родителей, фотогалереи, новости. Информация постоянно обновляется. | |
Методическое объединение преподавателей-педсовет, методика, учеба, образование - pedsovet. info : программы,рефераты, информатика, задачи, решения info : программы,рефераты, информатика, задачи, решения
| |
Химия, решение задач всех разделов, также прорешены все методички Шимановича И.Л., цены самые низкие. |
|
РГР по Теоретической Механике, Физике — Яблонский А.А, Мещерский И.В., Тарг С.М., Чертов А.Г, Воробьёв А.А, Волькенштейн В.С, Иродов Е.И, Трофимова Т.И., шпаргалки, лекции, учебники, много бесплатного. |
|
Макроэкономика-изучение основ дисциплины по лучшим лекциям и книгам. |
|
Оказания помощи школьникам м студентам в изучении физики и других предметов. На этом ресурсе размещены различные материалы: учебники, задачники, лекции, другие учебные пособия. |
|
Курсовые работы, дипломы, чертежи по процессам и аппаратам (ПАХТ) |
|
Диплом & Реферат- дипломные работы на заказ без предоплатыпо экономике, праву, туризму, общественным и гуманитарным предметам в соответствии с требованиями заказчика.
|
|
Рефераты, контрольные, курсовые, дипломные, шпаргалки |
Заказать РГР по сопромату, термеху в Санкт-Петербурге
Расчетно-графическая работа – это удел, как правило, студентов технических специальностей. Для того, чтобы качественно выполнить РГР, необходимо обладать хорошим логическим и образным мышлением, а также уметь применять теоретические знания на практике. Само название говорит о сути работы – необходимо произвести расчеты и в соответствии с полученными сведениями отобразить полученный результат графически. Многие студенты тратят на подобное задание уйму времени, пытаясь выполнить его самостоятельно, однако не всегда добиваются хорошего результата, и как итог – преподаватель отправляет переделывать задание заново. Если не уверенны в собственных силах и не имеете богатого опыта выполнения РГР, задумайтесь, стоит ли тратить свое драгоценное время и силы на попытку, которая с большой вероятностью потерпит фиаско?
Сколько стоит написание РГР на заказ?
Стоимость РГР в Санкт-Петербурге — от 99 ₽. Сроки выполнения — от 1 дня.
Сроки выполнения — от 1 дня.
Бесплатные доработки. Прохождение проверки на уникальность. Гарантия написания или возврат денег.
Наша компания предлагает студентам всех направлений и специальностей быстрое и простое решение проблем, связанных с выполнением работ различного рода, включая и РГР на заказ. Обеспечиваем клиентам максимально удобные и комфортные условия сотрудничества, именно поэтому каждому прикрепляется личный менеджер, который ведет заявку от ее оформления до закрытия. С ним можно связаться практически в любое удобное время и узнать на какой стадии выполнения находится заявка, а также получить все необходимые разъяснения и консультации по интересующим вопросам.
Можете всего несколькими щелчками мышки заказать выполнение ргр по сопромату, термеху, электротехнике, строительной механике, статистике, технической механике, термодинамике, физике и и др. Материалы, предоставленные сотрудниками, будут выполнены в четком соответствии с заявленными требованиями и предоставленными методическими рекомендациями. Все расчеты будут произведены и описаны максимально подробно для того, чтобы в случае, если преподаватель сочтет нужным уточнить какие-либо детали, без труда смогли бы ответить на все его вопросы, тем самым убедив в том, что заслуживаете действительно высокого балла за работу.
Все расчеты будут произведены и описаны максимально подробно для того, чтобы в случае, если преподаватель сочтет нужным уточнить какие-либо детали, без труда смогли бы ответить на все его вопросы, тем самым убедив в том, что заслуживаете действительно высокого балла за работу.
Широкий выбор способов оплаты – также большое преимущество, ведь в нашем случае клиент сам решает, каким именно образом он оплатит услуги. Заключение договора с каждым из наших клиентов и наличие чеков об оплате придают уверенности в солидности фирмы и ее ответственности перед клиентом. К тому же, обеспечиваем полное сопровождение заявки вплоть до ее защиты, это значит, что даже в том случае, когда полностью оплатили работу и получили ее на руки, при возникновении необходимости ее доработки, ни в коем случае не отказываемся от задания и вносим необходимые коррективы совершенно бесплатно. Узнать стоимость необходимого задания можно оформив заявку или связавшись с менеджером по телефону, указанному на главной странице сайта.
Относительная скорость роста (RGR) ПЭГ-чувствительных и ПЭГ-резистентных клеток…
Контекст 1
… отсутствие ПЭГ, ПЭГ-чувствительные и ПЭГ-резистентные каллусы демонстрировали одинаковый RGR (рис. 2). ). Рост ПЭГ-чувствительной клеточной линии в стандартной среде LS явно ингибировался ПЭГ, в то время как рост устойчивой клеточной линии не влиял. Добавление 50 мМ NaCl в культуральную среду, содержащую ПЭГ, явно улучшало RGR линии клеток, чувствительных к ПЭГ, но не влияло на показатель каллусов, устойчивых к ПЭГ.Когда клеточные линии содержались в среде, не содержащей Na + (LS-Na + ) в отсутствие PEG, RGR подвергался лишь незначительному влиянию. Напротив, при добавлении ПЭГ в среду, не содержащую Na +, обе клеточные линии были сильно затронуты, и не было зарегистрировано различий между чувствительными к ПЭГ и резистентными …
Контекст 2
… A и E были оценены в условиях насыщения при высоком уровне CO 2 и низком уровне O 2 . На растения воздействовали питательным раствором, содержащим 0% или 15% ПЭГ в присутствии 0 мМ или 50 мМ NaCl.Измерения проводились через 6 дней лечения. Устьичную проводимость количественно определяли порометрией независимо от измерений А. Каждое значение представляет среднее значение 6SE (n=12). Значения, имеющие общую букву в каждом столбце (параметр), существенно не различаются при P <0,05. результаты уже были получены для этого вида (Bajji et al., 1998) и для других видов рода Atriplex (Jeschke and Stelter, 1983; Matoh et al., 1986; Mahmood and Malik, 1987). Эта работа, однако, в основном демонстрирует, что низкие уровни солености могут также улучшать способность растений справляться с водным стрессом, вызванным ПЭГ.Действительно, рост ( рис. 1 ), OA (таблица 1) и A (таблица 3) были явно выше у растений, обработанных ПЭГ, одновременно подвергавшихся воздействию 50 мМ NaCl, по сравнению с растениями, подвергнутыми воздействию ПЭГ в отсутствие соли. Наоборот, gs и E (таблица 3) явно снижались у растений, обработанных ПЭГ, при воздействии NaCl.
На растения воздействовали питательным раствором, содержащим 0% или 15% ПЭГ в присутствии 0 мМ или 50 мМ NaCl.Измерения проводились через 6 дней лечения. Устьичную проводимость количественно определяли порометрией независимо от измерений А. Каждое значение представляет среднее значение 6SE (n=12). Значения, имеющие общую букву в каждом столбце (параметр), существенно не различаются при P <0,05. результаты уже были получены для этого вида (Bajji et al., 1998) и для других видов рода Atriplex (Jeschke and Stelter, 1983; Matoh et al., 1986; Mahmood and Malik, 1987). Эта работа, однако, в основном демонстрирует, что низкие уровни солености могут также улучшать способность растений справляться с водным стрессом, вызванным ПЭГ.Действительно, рост ( рис. 1 ), OA (таблица 1) и A (таблица 3) были явно выше у растений, обработанных ПЭГ, одновременно подвергавшихся воздействию 50 мМ NaCl, по сравнению с растениями, подвергнутыми воздействию ПЭГ в отсутствие соли. Наоборот, gs и E (таблица 3) явно снижались у растений, обработанных ПЭГ, при воздействии NaCl. Результаты, полученные с выбранными клеточными линиями, позволяют предположить, что положительный эффект NaCl может быть обусловлен Na+. На стандартной среде LS устойчивые к ПЭГ каллусы накапливали больше Na+, чем чувствительные к ПЭГ (рис. 4), в то время как для хлорида различий не наблюдалось.Однако при воздействии ПЭГ на обе клеточные линии на среде без Na + различия между клеточными линиями больше не регистрировались, а «устойчивые» каллусы не могли расти (рис. 2) и эффективно выполнять ОА (табл. 1). ) в присутствии ПЭГ. Напротив, когда 50 мМ NaCl добавляли в среду LS, чувствительная к ПЭГ клеточная линия была способна накапливать количество Na +, аналогичное таковому у устойчивой к ПЭГ, и обе линии демонстрировали сходные характеристики с точки зрения роста и ОА. Таким образом, эти данные свидетельствуют о том, что Na + участвует в устойчивости к водному стрессу, индуцированному ПЭГ, и что чувствительная клеточная линия была менее эффективна, чем резистентная, в поглощении Na + из внешней среды, содержащей низкие концентрации Na +.
Результаты, полученные с выбранными клеточными линиями, позволяют предположить, что положительный эффект NaCl может быть обусловлен Na+. На стандартной среде LS устойчивые к ПЭГ каллусы накапливали больше Na+, чем чувствительные к ПЭГ (рис. 4), в то время как для хлорида различий не наблюдалось.Однако при воздействии ПЭГ на обе клеточные линии на среде без Na + различия между клеточными линиями больше не регистрировались, а «устойчивые» каллусы не могли расти (рис. 2) и эффективно выполнять ОА (табл. 1). ) в присутствии ПЭГ. Напротив, когда 50 мМ NaCl добавляли в среду LS, чувствительная к ПЭГ клеточная линия была способна накапливать количество Na +, аналогичное таковому у устойчивой к ПЭГ, и обе линии демонстрировали сходные характеристики с точки зрения роста и ОА. Таким образом, эти данные свидетельствуют о том, что Na + участвует в устойчивости к водному стрессу, индуцированному ПЭГ, и что чувствительная клеточная линия была менее эффективна, чем резистентная, в поглощении Na + из внешней среды, содержащей низкие концентрации Na +. Различия между клеточными линиями исчезали, когда более высокие внешние концентрации Na+ были …
Различия между клеточными линиями исчезали, когда более высокие внешние концентрации Na+ были …
Фактические концентрации цитокинина в тканях растений как показатель солеустойчивости зерновых по JSTOR
Абстрактный Скорость роста корней и побегов выращенных на гидропонике растений солеустойчивых сортов ячменя быстро снижалась после воздействия 65 Мм⁻³ хлорида натрия. Это снижение скорости роста, как правило, было временным и сопровождалось пониженным отношением побегов к корням.В то же время происходило замедление роста, также быстро снижались концентрации цитокининов в ткани корней и побегов, определяемые с помощью иммуноферментного анализа. Добавление 5·10⁻⁹ Mm⁻3 бензиладенина (BA) замедляло снижение скорости роста, отношения побегов к корням и концентрации внутренних цитокининов. Интенсивность роста корней и побегов одинаково выращенных растений солечувствительных сортов ячменя сохраняла свои темпы роста и соотношение побегов и корней после воздействия NaCl не менее 10 сут в сочетании с неизмененным уровнем эндогенных цитокининов. Примерно через 14 дней NaCl вызывал резкое снижение значений RGR, но не отношения побегов к корням. Добавление бензиладенина в некоторой степени ингибировало рост и повышало концентрацию цитокинина. Был сделан вывод, что концентрации цитокининов, измеренные непосредственно после воздействия NaCl, представляют собой многообещающий метод скрининга на солеустойчивость. Добавление БА привело к тому, что солеустойчивые сорта стали вести себя как солеустойчивые сорта.
Примерно через 14 дней NaCl вызывал резкое снижение значений RGR, но не отношения побегов к корням. Добавление бензиладенина в некоторой степени ингибировало рост и повышало концентрацию цитокинина. Был сделан вывод, что концентрации цитокининов, измеренные непосредственно после воздействия NaCl, представляют собой многообещающий метод скрининга на солеустойчивость. Добавление БА привело к тому, что солеустойчивые сорта стали вести себя как солеустойчивые сорта.
Plant and Soil публикует оригинальные статьи и обзорные статьи, исследующие взаимодействие биологии растений и наук о почве и предлагающие четкий механистический компонент.Сюда входят как фундаментальные, так и прикладные аспекты минерального питания, отношений растений и воды, симбиотических и патогенных взаимодействий растений и микробов, анатомии и морфологии корней, биологии почв, экологии, агрохимии и агрофизики. Статьи, посвященные основным молекулярным или математическим компонентам, также входят в сферу охвата журнала.
Springer — одно из ведущих международных научных издательств, выпускающее более 1200 журналов и более 3000 новых книг ежегодно, охватывающих широкий круг предметов, включая биомедицину и науки о жизни, клиническую медицину, физика, инженерия, математика, информатика и экономика.
Влияние устойчивости к гербицидам, обуславливающей мутации гена АССазы Ile-1781-Leu и Asp-2078-Gly, на кинетику АССазы и признаки роста Lolium Ridium | Журнал экспериментальной ботаники
Аннотация
Скорость эволюции устойчивости растений к гербицидам зависит от признаков приспособленности, придаваемых аллелями как в присутствии, так и в отсутствие (цена устойчивости) селекции гербицидов. Здесь было исследовано влияние двух спонтанных гомозиготных мутаций Lolium Rigidum , придающих резистентность к сайту-мишени (Ile-1781-Leu, Asp-2078-Gly), как на активность АССазы, так и на различные признаки роста растений. Были оценены относительная скорость роста (RGR) и компоненты (чистая скорость ассимиляции, соотношение площади листьев), распределение ресурсов по различным органам и реакция роста в конкуренции с урожаем пшеницы. В отличие от растений, несущих мутацию устойчивости к Ile-1781-Leu, растения, гомозиготные по мутации Asp-2078-Gly, демонстрировали значительно более низкий RGR (30%), что выражалось в более низком распределении биомассы корням, побегам и листьям и плохой реакции. разводить конкуренцию. Как незначительное, так и значительное снижение роста, связанное, соответственно, с мутациями устойчивости к Ile-1781-Leu и Asp-2078-Gly, коррелирует с их влиянием на активность АССазы.В то время как мутация Ile-1781-Leu не проявляла плейотропных эффектов на кинетику АССазы, мутация Asp-2078-Gly приводила к значительному снижению активности АССазы. Признаки нарушения роста обсуждаются в контексте затрат на устойчивость и влияния каждого аллеля устойчивости на активность АССазы. Аналогичные эффекты этих двух конкретных мутаций ACCase на активность ACCase Alopecurus myosuroides также были подтверждены.
Были оценены относительная скорость роста (RGR) и компоненты (чистая скорость ассимиляции, соотношение площади листьев), распределение ресурсов по различным органам и реакция роста в конкуренции с урожаем пшеницы. В отличие от растений, несущих мутацию устойчивости к Ile-1781-Leu, растения, гомозиготные по мутации Asp-2078-Gly, демонстрировали значительно более низкий RGR (30%), что выражалось в более низком распределении биомассы корням, побегам и листьям и плохой реакции. разводить конкуренцию. Как незначительное, так и значительное снижение роста, связанное, соответственно, с мутациями устойчивости к Ile-1781-Leu и Asp-2078-Gly, коррелирует с их влиянием на активность АССазы.В то время как мутация Ile-1781-Leu не проявляла плейотропных эффектов на кинетику АССазы, мутация Asp-2078-Gly приводила к значительному снижению активности АССазы. Признаки нарушения роста обсуждаются в контексте затрат на устойчивость и влияния каждого аллеля устойчивости на активность АССазы. Аналогичные эффекты этих двух конкретных мутаций ACCase на активность ACCase Alopecurus myosuroides также были подтверждены.
Введение
Ацетил-коэнзим А-карбоксилаза (ACCase) является ключевым растительным ферментом в биосинтезе липидов и является мишенью гербицидов, ингибирующих ACCase (далее именуемых гербицидами ACCase).Был идентифицирован ряд развившихся одиночных мутаций гена ACCase , которые придают устойчивость к гербицидам ACCase у видов злаковых сорняков (рассмотрено в Délye, 2005; Powles and Yu, 2010; Kaundun, 2014). У Lolium Rigidum , самого распространенного сорняка в сельском хозяйстве Австралии, эти мутации гена ACCase приводят к аминокислотным заменам ACCase, включая Ile-1781-Leu/Val, Trp-1999-Cys/Leu, Ile-2041-Asn. /Asp/Thr/Val, Asp-2078-Gly, Cys-2088-Arg/Phe и Gly-2096-Ala (Yu et al., 2007b; Мэлоун и др. , 2013). Молекулярный и исследовательский анализы показали, что частота и распределение этих эволюционировавших мутантных аллелей ACCase различаются среди популяций (Yu et al. , 2007b; Malone et al. , 2013; Vila-Aiub et al. , 2009b; Кешткар и др. , 2015).
, 2007b; Malone et al. , 2013; Vila-Aiub et al. , 2009b; Кешткар и др. , 2015).
Кристаллические структуры домена карбокситрансферазы ACCase дрожжевой ACCase показали, что гербициды ACCase связаны в активном центре карбокситрансферазы (Zhang et al., 2004). Таким образом, специфические мутации гена ACCase , придавая устойчивость к гербицидам, могут также оказывать влияние на активность ACCase (Délye et al. , 2005). Например, мутация Asp-2078-Gly придает высокий уровень устойчивости ( I 50 , по сравнению с чувствительным аналогом) к диклофоп-метилу, галоксифопу и тралкоксидиму, а также специфическую активность АССазы (в отсутствие обработки гербицидами) значительно снижается (Ю и др., 2007b).
Мутации резистентности к гербицидам в месте-мишени могут повлечь за собой затраты на резистентность из-за нарушения каталитической способности фермента и/или снижения сродства к субстрату и/или изменения ингибирования по принципу обратной связи, приводящего к недостаточному или избыточному биосинтезу продукта (Purrington and Bergelson, 1999; Vila-Aiub et и др.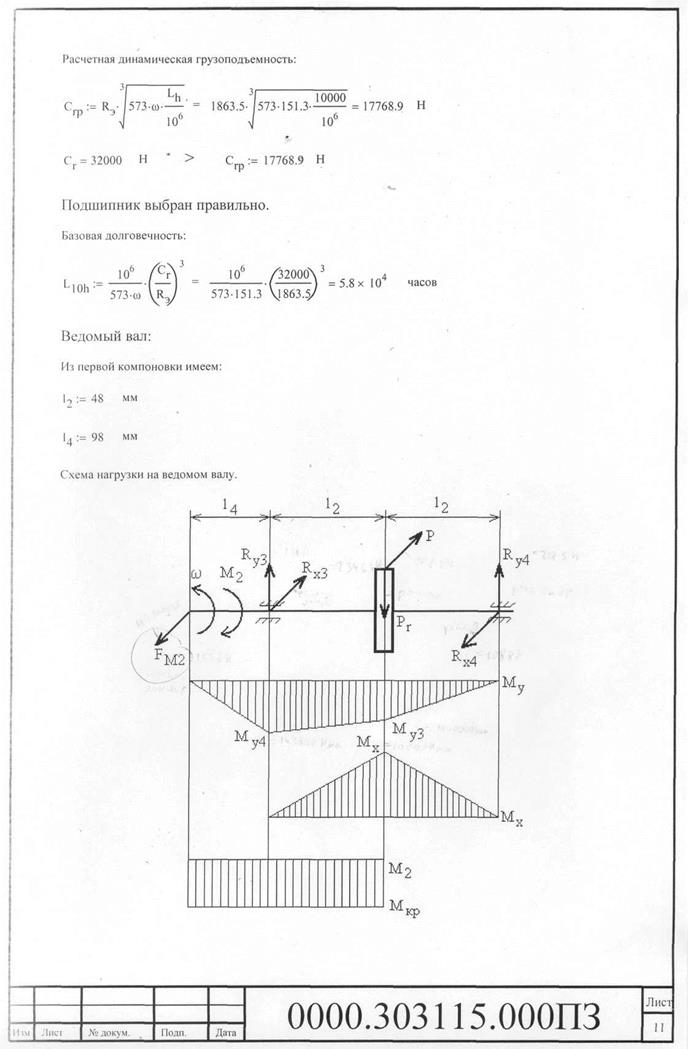 , 2009б; Ю и др. , 2010). В экологическом эволюционном контексте возможна быстрая эволюция устойчивости к гербицидам, если мутация гена обеспечивает значительный уровень устойчивости и при этом не показывает никаких или незначительных затрат на пригодность (величина таких затрат оценивается в отсутствие селекции гербицидов) (Вила-Аюб). и др., 2015). Наоборот, генетические признаки, обеспечивающие высокий или низкий уровень устойчивости и связанные с приспособленностью, скорее всего, эволюционируют относительно медленно.
, 2009б; Ю и др. , 2010). В экологическом эволюционном контексте возможна быстрая эволюция устойчивости к гербицидам, если мутация гена обеспечивает значительный уровень устойчивости и при этом не показывает никаких или незначительных затрат на пригодность (величина таких затрат оценивается в отсутствие селекции гербицидов) (Вила-Аюб). и др., 2015). Наоборот, генетические признаки, обеспечивающие высокий или низкий уровень устойчивости и связанные с приспособленностью, скорее всего, эволюционируют относительно медленно.
Здесь плейотропные эффекты мутаций Ile-1781-Leu и Asp-2078-Gly гена АССазы как на активность АССазы, так и на кинетику и связанные с приспособленностью признаки роста были оценены у L. Riginum . Это исследование обеспечивает лучшее понимание биохимической основы стоимости устойчивости и эволюционной динамики аллелей устойчивости к гербицидам ACCase у L.ригидум .
Материалы и методы
Растительный материал и мутации гена устойчивости к гербицидам ACCase
Ряд выведенных в полевых условиях устойчивых к ACCase-гербицидам популяций L. hardum , собранных в пшеничном поясе Западной Австралии (Owen et al. , 2007), был подвергнут детальной молекулярной характеристике, что позволило идентифицировать популяции, устойчивые к ACCase-гербицидам благодаря специфическому Мутации гена ACCase (Yu et al., 2007b). Были созданы и полностью охарактеризованы очищенные популяции, каждая из которых была гомозиготной ( RR ) по специфическим мутациям устойчивости к гербицидам ACCase Ile-1781-Leu и Asp-2078-Gly (Yu et al. , 2007b). Это было достигнуто путем выявления (с помощью секвенирования и анализа маркеров на основе ПЦР) растений, гомозиготных по конкретной мутации, и выращивания этих гомозиготных растений до зрелости и обеспечения перекрестного опыления в защищенных от пыльцы клетках для получения семян для одного поколения.Гомозиготность растений-потомков по специфической мутации гена ACCase в каждой очищенной популяции подтверждали с помощью маркерного анализа на основе ПЦР, как описано в Yu et al.
hardum , собранных в пшеничном поясе Западной Австралии (Owen et al. , 2007), был подвергнут детальной молекулярной характеристике, что позволило идентифицировать популяции, устойчивые к ACCase-гербицидам благодаря специфическому Мутации гена ACCase (Yu et al., 2007b). Были созданы и полностью охарактеризованы очищенные популяции, каждая из которых была гомозиготной ( RR ) по специфическим мутациям устойчивости к гербицидам ACCase Ile-1781-Leu и Asp-2078-Gly (Yu et al. , 2007b). Это было достигнуто путем выявления (с помощью секвенирования и анализа маркеров на основе ПЦР) растений, гомозиготных по конкретной мутации, и выращивания этих гомозиготных растений до зрелости и обеспечения перекрестного опыления в защищенных от пыльцы клетках для получения семян для одного поколения.Гомозиготность растений-потомков по специфической мутации гена ACCase в каждой очищенной популяции подтверждали с помощью маркерного анализа на основе ПЦР, как описано в Yu et al. (2007b). Потомство массового скрещивания двух очищенных устойчивых популяций использовали во всех последующих экспериментах. Этот экспериментальный подход исключает любые смешанные эффекты потенциальных различий в затратах на устойчивость, связанных с преобладанием стоимости ( RR против RS ) (Roux et al., 2004). Четыре чувствительные к гербицидам популяции L. hardum (h4/6, h5/6, h5/33 и VLR1, далее именуемые S 1 , S 2 , S 3 и S 4 соответственно ), проявляющие чувствительность к ACCase (Yu et al. , 2007b; Yu et al. , 2009), использовали в качестве контроля дикого типа, чтобы свести к минимуму различия в генетическом фоне между устойчивыми к гербицидам ACCase и различными восприимчивыми популяциями (Vila-Aiub). и др. , 2009b, Vila-Aiub и др., 2011). Этот экспериментальный подход предполагает, что статистически значимая разница в средних значениях признаков между сравниваемыми устойчивыми и восприимчивыми популяциями указывает на то, что эти различия, вероятно, вызваны плейотропными эффектами гена(ов) устойчивости (Cousens et al.
(2007b). Потомство массового скрещивания двух очищенных устойчивых популяций использовали во всех последующих экспериментах. Этот экспериментальный подход исключает любые смешанные эффекты потенциальных различий в затратах на устойчивость, связанных с преобладанием стоимости ( RR против RS ) (Roux et al., 2004). Четыре чувствительные к гербицидам популяции L. hardum (h4/6, h5/6, h5/33 и VLR1, далее именуемые S 1 , S 2 , S 3 и S 4 соответственно ), проявляющие чувствительность к ACCase (Yu et al. , 2007b; Yu et al. , 2009), использовали в качестве контроля дикого типа, чтобы свести к минимуму различия в генетическом фоне между устойчивыми к гербицидам ACCase и различными восприимчивыми популяциями (Vila-Aiub). и др. , 2009b, Vila-Aiub и др., 2011). Этот экспериментальный подход предполагает, что статистически значимая разница в средних значениях признаков между сравниваемыми устойчивыми и восприимчивыми популяциями указывает на то, что эти различия, вероятно, вызваны плейотропными эффектами гена(ов) устойчивости (Cousens et al. , 1997; Strauss et al. , 2002). Методологические подходы, использованные в этих исследованиях, позволяют проводить независимое сравнение каждой популяции, устойчивой к гербицидам ACCase, со всеми популяциями, чувствительными к гербицидам ACCase.Информация о шести популяциях L. hardum , использованных в этом исследовании, приведена в дополнительной таблице S1.
, 1997; Strauss et al. , 2002). Методологические подходы, использованные в этих исследованиях, позволяют проводить независимое сравнение каждой популяции, устойчивой к гербицидам ACCase, со всеми популяциями, чувствительными к гербицидам ACCase.Информация о шести популяциях L. hardum , использованных в этом исследовании, приведена в дополнительной таблице S1.
Чтобы сопоставить влияние конкретных мутаций устойчивости к гербицидам ACCase (Ile-1781-Leu или Asp-2078-Gly) на выражение стоимости устойчивости как на уровне всего растения, так и на уровне фермента, активности ACCase и кинетики, связанной с каждой устойчивостью мутация была оценена. Особи из восприимчивой популяции S 4 и две другие популяции L. Rigidum , собранные из систем посева WA (WALR60, WALR70) с известными восприимчивыми последовательностями ACCase, служили в качестве контроля дикого типа для измерения активности и кинетики ACCase (Yu et al. ., 2007а; Ю и др. , 2009 г.). Поскольку затраты на приспособляемость, связанные с мутацией Asp-2078-Gly, но не с мутацией Ile-1781-Leu, ранее сообщались у Alopecurus myosuroides (Menchari et al. , 2008), влияние этих двух мутаций устойчивости на Поэтому активность АССазы также оценивали у A. myosuroides . Очищенная популяция A. myosuroides , гомозиготная по мутации Asp-2078-Gly (Délye et al. , 2003; Délye et al., 2005) и полевая популяция, содержащая 92% растений, гомозиготных по мутации Ile-1781-Leu (Délye, личное сообщение). Две чувствительные к ACCase гербицидам популяции A. myosuroides использовали в качестве контроля дикого типа.
, 2009 г.). Поскольку затраты на приспособляемость, связанные с мутацией Asp-2078-Gly, но не с мутацией Ile-1781-Leu, ранее сообщались у Alopecurus myosuroides (Menchari et al. , 2008), влияние этих двух мутаций устойчивости на Поэтому активность АССазы также оценивали у A. myosuroides . Очищенная популяция A. myosuroides , гомозиготная по мутации Asp-2078-Gly (Délye et al. , 2003; Délye et al., 2005) и полевая популяция, содержащая 92% растений, гомозиготных по мутации Ile-1781-Leu (Délye, личное сообщение). Две чувствительные к ACCase гербицидам популяции A. myosuroides использовали в качестве контроля дикого типа.
Активность и кинетика ACCase, связанные со специфическими мутациями устойчивости к гербицидам ACCase
Особи, гомозиготные по каждой конкретной мутации устойчивости к ACCase (Ile-1781-Leu или Asp-2078-Gly), и восприимчивые особи выращивались в тепличных условиях (подробности см. ниже).На стадии трех-четырех листьев надземный листовой материал (около 3 г) собирали на уровне почвы из каждой популяции (не менее 30–40 саженцев за урожай), быстро замораживали в жидком азоте и хранили при температуре — 80°С. Анализ ACCase in vitro проводили в соответствии со способом Yu et al. (2004 г.) с модификациями. Замороженный материал измельчали в мелкий порошок с помощью ступки и пестика в жидком азоте и гомогенизировали в 10 мл буфера для экстракции, содержащего 100 мМ трис (pH 8.0), 1 мМ ЭДТА, 10% (об./об.) глицерина, 2 мМ изоаскорбиновой кислоты, 1 мМ PMSF, 0,5% ПВП-40, 0,5% нерастворимого ПВП и 20 мМ ДТТ. Гомогенат центрифугировали при 27000 g в течение 15 мин. Супернатант доводили до 40% насыщения (NH 4 ) 2 SO 4 добавлением по каплям насыщенного раствора (NH 4 ) 2 SO 4 и перемешивали в течение 10 мин. Раствор центрифугировали при 27000 g в течение 20 мин.
ниже).На стадии трех-четырех листьев надземный листовой материал (около 3 г) собирали на уровне почвы из каждой популяции (не менее 30–40 саженцев за урожай), быстро замораживали в жидком азоте и хранили при температуре — 80°С. Анализ ACCase in vitro проводили в соответствии со способом Yu et al. (2004 г.) с модификациями. Замороженный материал измельчали в мелкий порошок с помощью ступки и пестика в жидком азоте и гомогенизировали в 10 мл буфера для экстракции, содержащего 100 мМ трис (pH 8.0), 1 мМ ЭДТА, 10% (об./об.) глицерина, 2 мМ изоаскорбиновой кислоты, 1 мМ PMSF, 0,5% ПВП-40, 0,5% нерастворимого ПВП и 20 мМ ДТТ. Гомогенат центрифугировали при 27000 g в течение 15 мин. Супернатант доводили до 40% насыщения (NH 4 ) 2 SO 4 добавлением по каплям насыщенного раствора (NH 4 ) 2 SO 4 и перемешивали в течение 10 мин. Раствор центрифугировали при 27000 g в течение 20 мин. Осадок ресуспендировали в 1,5 мл элюирующего буфера (50 мМ трицина, рН 8.0, 2,5 мМ MgCl 2 , 50 мМ KCl, 1 мМ DTT) и обессоливали на колонке Sephadex G-25, предварительно уравновешенной элюирующим буфером. Определяли концентрацию белка в каждом обессоленном образце (Bradford, 1976), и образец немедленно использовали для анализа.
Осадок ресуспендировали в 1,5 мл элюирующего буфера (50 мМ трицина, рН 8.0, 2,5 мМ MgCl 2 , 50 мМ KCl, 1 мМ DTT) и обессоливали на колонке Sephadex G-25, предварительно уравновешенной элюирующим буфером. Определяли концентрацию белка в каждом обессоленном образце (Bradford, 1976), и образец немедленно использовали для анализа.
Активность АССазы определяли путем количественного определения включения NaH 14 CO 3 в кислотоустойчивый продукт малонил-КоА. Экстракт фермента инкубировали при 30°C в реакционной смеси, содержащей 10 мМ Tricine-KOH, pH 8.3, 5 мм ATP, 10 мм MGCL 2 , 0,1% BSA, 2,5 мм DTT и 10 мм Nahco 3 (дополнены в среднем в среднем 24 KBQ Nah 14 CO 3 , 2.18 GBQ MMOL -1 ) . Реакцию начинали добавлением ацетил-КоА в конечной концентрации 0,5 мМ и останавливали через 10 минут добавлением концентрированной HCl. Анализы без ацетил-КоА использовали в качестве контроля. Кислотостабильную радиоактивность измеряли сцинтилляционным счетчиком, а активность АССазы рассчитывали методом изотопного разбавления. Для кинетического измерения ACCase (проведенного только для L. Ridum ) использовали 40–120 мкг белка, поскольку этот уровень белка катализирует линейную скорость образования малонил-КоА в этих экспериментальных условиях. Самая высокая концентрация 1,5, 3 и 20 мМ ацетил-КоА, АТФ и HCO 3 соответственно использовалась при определении значений Km . Общий белок нормализовали до 100 мкг для всех образцов, чтобы измерить специфичную для АССазы активность.
Для кинетического измерения ACCase (проведенного только для L. Ridum ) использовали 40–120 мкг белка, поскольку этот уровень белка катализирует линейную скорость образования малонил-КоА в этих экспериментальных условиях. Самая высокая концентрация 1,5, 3 и 20 мМ ацетил-КоА, АТФ и HCO 3 соответственно использовалась при определении значений Km . Общий белок нормализовали до 100 мкг для всех образцов, чтобы измерить специфичную для АССазы активность.
Значения Km рассчитаны с использованием нелинейного регрессионного анализа путем подгонки данных к уравнению Михаэлиса–Ментен пирувата, ν — скорость реакции при любой концентрации пирувата, V — максимальная скорость реакции ( Vmax ).Каждый анализ содержал две технические повторности, и для каждого набора анализов использовали четыре независимых ферментных экстракта. Данные подвергали дисперсионному анализу с использованием программного обеспечения SAS (версия 9.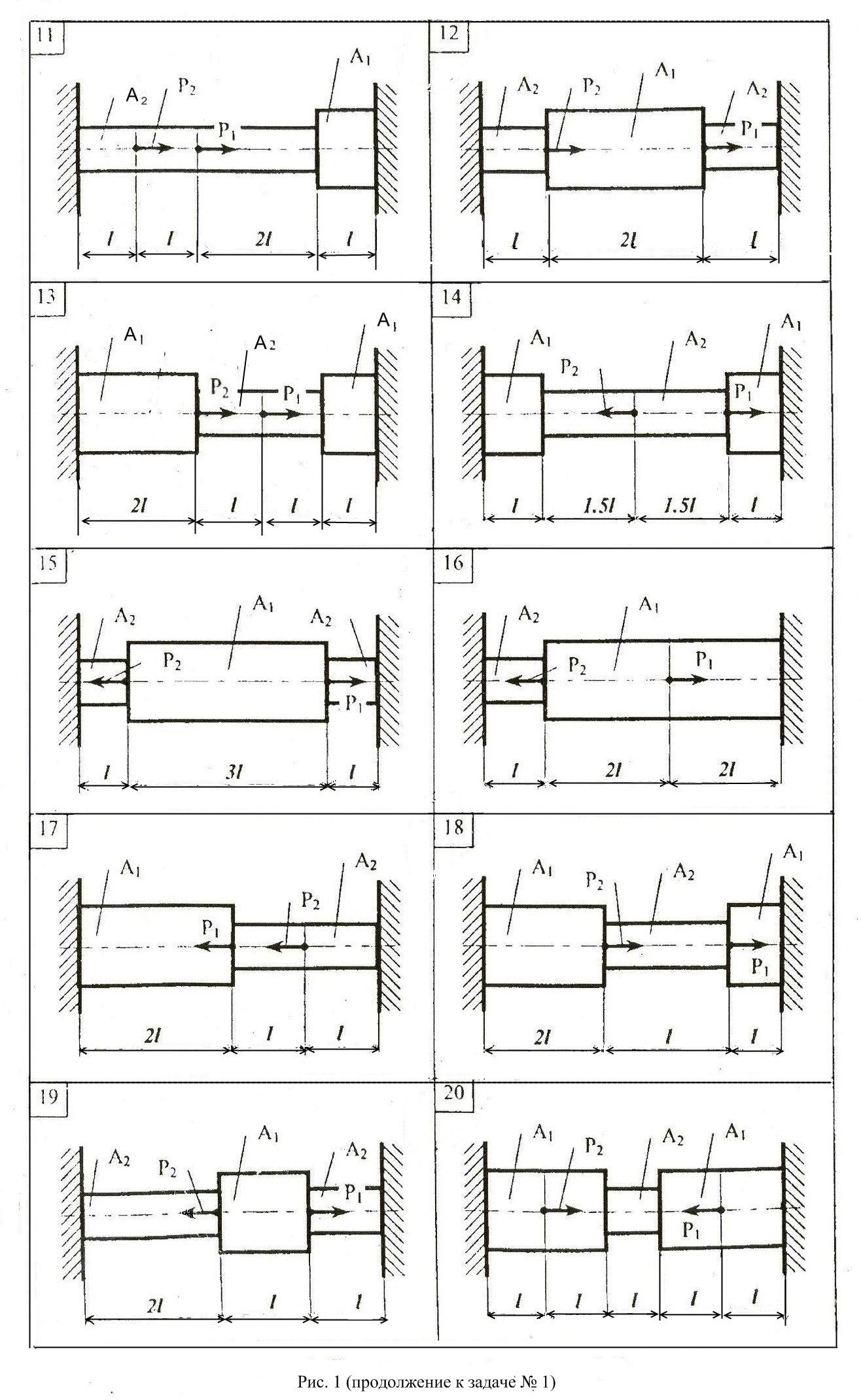 3, Cary, NC, SAS Institute Inc., 2002–2010 гг.). Средние значения были разделены с использованием защищенного критерия наименьшего значимого различия (LSD) Фишера при уровне вероятности 5%.
3, Cary, NC, SAS Institute Inc., 2002–2010 гг.). Средние значения были разделены с использованием защищенного критерия наименьшего значимого различия (LSD) Фишера при уровне вероятности 5%.
Стоимость резистентности, связанная со специфическими мутациями
ACCase гена резистентности Эксперименты, предназначенные для оценки роста обоих изолированных растений (т.е. отсутствие конкуренции) и в условиях межвидовой конкуренции проводились дважды. Относительная скорость роста (RGR) и компоненты [чистая скорость ассимиляции (NAR), коэффициент площади листьев (LAR)], а также распределение ресурсов на корни, стебли и площадь листьев оценивались у изолированных растений, растущих без конкуренции. Конкурентные реакции L.rigidum , экспрессирующих специфические мутации устойчивости к АССазе к гербицидам (Ile-1781-Leu или Asp-2078-Gly) и чувствительного к дикому типу, оценивали в условиях конкуренции с пшеницей ( Triticum aestivum ). Как RGR, так и реакция на конкуренцию за ресурсы являются полезными экофизиологическими параметрами для обозначения затрат на устойчивость к гербицидам, поскольку вариации этих признаков положительно коррелируют с вариациями конкурентоспособности растений, способности к укоренению и плодовитости (Grime and Hunt, 1975; Goldberg, 1990; Weiner, 2004; Vila-Aiub и др. , 2005).
Как RGR, так и реакция на конкуренцию за ресурсы являются полезными экофизиологическими параметрами для обозначения затрат на устойчивость к гербицидам, поскольку вариации этих признаков положительно коррелируют с вариациями конкурентоспособности растений, способности к укоренению и плодовитости (Grime and Hunt, 1975; Goldberg, 1990; Weiner, 2004; Vila-Aiub и др. , 2005).
Выращивание изолированных растений без конкуренции
Семена гомозиготных устойчивых к гербицидам АССазы мутантов (Ile-1781-Leu, Asp-2078-Gly) и восприимчивых генотипов дикого типа (от S 1 до S 4 ) проращивали на 0.7% (масса/объем) агар (12 часов на свету при 25°C, 12 часов в темноте при 15°C). Через 4 дня отдельные сеянцы (высотой 2 см) пересаживали в отдельные горшки (диаметром 9 см, высотой 13 см), содержащие стандартную почвенную смесь (50% торфяного мха и 50% речного песка). Растения выращивали в теплице, расположенной в полностью случайном порядке. Горшки регулярно переставляли, чтобы рандомизировать любые различия в окружающей среде в теплице. Средние температурные условия выращивания колебались от 20°C (днем) до 15°C (ночью), что близко к оптимальному для этого вида.Урожай собирали через 10 и 24 дня после высадки рассады. При каждом урожае оценивали надземную и корневую биомассу и площадь листьев. Растения извлекали из почвы и промывали корни водопроводной водой. Площадь листьев на одном растении определяли с помощью цифрового измерителя площади листьев (LI-3100; LiCor, Линкольн, Небраска, США). Надземный материал (побеги делили на листовой материал и стебли, включая листовую оболочку) и корни сушили в печи при 80°C в течение 72 часов и регистрировали сухую биомассу. RGR и его компоненты (NAR и LAR) рассчитывали для каждой комбинации обработок (генотип × урожай).Было 20 повторов на обработку (семь генотипов × два времени сбора урожая). Растения регулярно поливали и еженедельно удобряли жидким удобрением [N 19% (NH 2 15%, NH 4 1,9%, NO 3 2,1%), P 8%, K 16%, Mg 1,2%, S 3,8%, Fe 400 частей на миллион, Mn 200 частей на миллион, Zn 200 частей на миллион, Cu 100 частей на миллион, B 100 частей на миллион и Mo 10 частей на миллион].
Горшки регулярно переставляли, чтобы рандомизировать любые различия в окружающей среде в теплице. Средние температурные условия выращивания колебались от 20°C (днем) до 15°C (ночью), что близко к оптимальному для этого вида.Урожай собирали через 10 и 24 дня после высадки рассады. При каждом урожае оценивали надземную и корневую биомассу и площадь листьев. Растения извлекали из почвы и промывали корни водопроводной водой. Площадь листьев на одном растении определяли с помощью цифрового измерителя площади листьев (LI-3100; LiCor, Линкольн, Небраска, США). Надземный материал (побеги делили на листовой материал и стебли, включая листовую оболочку) и корни сушили в печи при 80°C в течение 72 часов и регистрировали сухую биомассу. RGR и его компоненты (NAR и LAR) рассчитывали для каждой комбинации обработок (генотип × урожай).Было 20 повторов на обработку (семь генотипов × два времени сбора урожая). Растения регулярно поливали и еженедельно удобряли жидким удобрением [N 19% (NH 2 15%, NH 4 1,9%, NO 3 2,1%), P 8%, K 16%, Mg 1,2%, S 3,8%, Fe 400 частей на миллион, Mn 200 частей на миллион, Zn 200 частей на миллион, Cu 100 частей на миллион, B 100 частей на миллион и Mo 10 частей на миллион].
Программное обеспечение (Hunt et al. , 2002) использовалось для расчета параметров роста, полученных в соответствии с классическим анализом роста (Hunt, 1982; Poorter and Nagel, 2000).Для расчета RGR использовалась несмещенная формула, предложенная Hoffmann and Poorter (2002). Дисперсия (σ 2 или V ), связанная с RGR, оценивалась по формуле Causton and Venus (1981). Степени свободы, связанные с RGR и его компонентами, составили n – 2, где n – общее количество растений, использованных в двух урожаях. Односторонний ANOVA был выполнен для сравнения RGR и его компонентов на предмет устойчивости к ACCase и восприимчивых генотипов L. hardum дикого типа.Апостериорный критерий Даннета использовали для сравнения средних значений генотипов устойчивости к гербицидам ACCase с восприимчивыми эталонными генотипами дикого типа (α = 5%).
Стоимость устойчивости, связанная с каждой из мутаций устойчивости к гербицидам ACCase, оценивалась в соответствии с Vila-Aiub et al. (2009b):
(2009b):
RC(R/S)(%)=[1−(rRrS)]*100
(1)где RC ( R/S ) представляет стоимость сопротивления ( RC ) устойчивого к гербицидам ( R ) генотипа относительно чувствительного к гербицидам ( S ) генотипа, rR обозначает ответ устойчивого генотипа R , а rS представляет собой ответ восприимчивого дикого типа генотип S .
Выращивание растений на условиях конкуренции
Для оценки реакции конкуренции за ресурсы у гомозиготных устойчивых к гербицидам мутантов АССазы (Ile-1781-Leu, Asp-2078-Gly) и восприимчивых генотипов дикого типа (от S 1 до S 4) применялся план эксперимента с целевым окружением. ), выращенный в конкуренции с пшеницей (дополнительный рисунок S1) (Gibson et al. , 1999). Конкурентная реакция на ресурсы окружающей среды связана со способностью растения сохраняться независимо от присутствия конкурента (пшеницы). Таким образом, вегетативную активность целевых растений (т.е. генотип ACCase RR или SS ) оценивали при увеличении плотности и биомассы соседних растений пшеницы (Weiner, 1982; Goldberg and Werner, 1983).
Таким образом, вегетативную активность целевых растений (т.е. генотип ACCase RR или SS ) оценивали при увеличении плотности и биомассы соседних растений пшеницы (Weiner, 1982; Goldberg and Werner, 1983).
Оценку конкурентных ответов целевых растений ACCase RR и SS проводили в условиях асимметричной по размеру конкуренции со стороны пшеницы (Goldberg, 1990). Пшеница (сорт Wyalkatchem) была посеяна в горшки (диаметр 25 см × высота 23 см), содержащие почвенную смесь (50% торфяного мха и 50% речного песка) в соответствии со схемами посадки (дополнительный рис.С1). Семена одинаковой массы генотипов ACCase RR и SS проращивали, как описано ранее. Когда пшеница была на стадии трех распустившихся листьев (высота 15 см), сеянцы L. hardum на стадии одного листа (высотой 2 см) пересаживали в горшки, содержащие пшеницу. Удобрение с медленным высвобождением (Macrocote Blue Plus) (12 г/горшок) [вес/вес N 16 %, (NH 2 8,4 %, NH 4 6,45 %, NO 3 1,47 %), P 4 %, K 10%, S 5%, Mg 0,63%, Fe 0,20%, Cu 0,03%, Zn 0.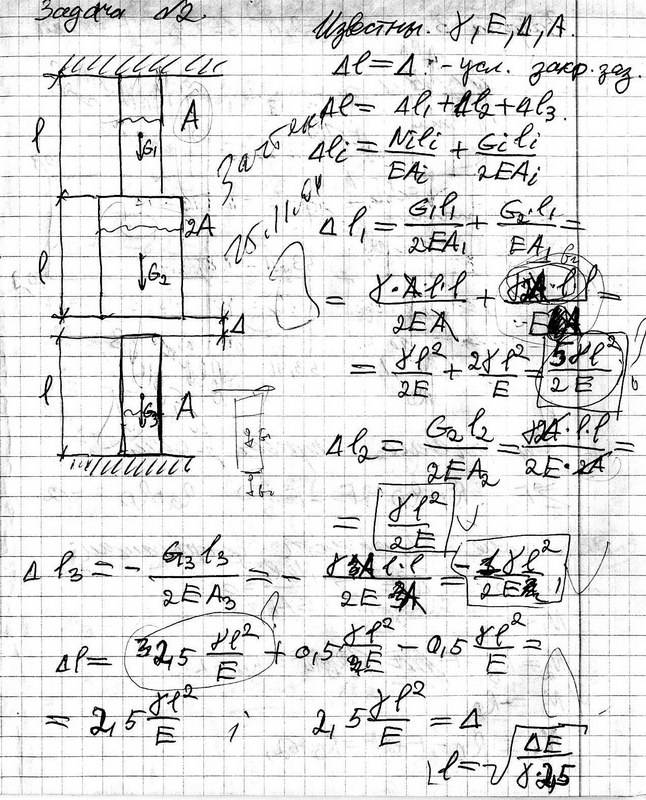 03% и Mn 0,08%] и жидкие удобрения [N 19% (NH 2 15%, NH 4 1,9%, NO 3 2,1%), P 8%, K 16%, Mg 1,2%, S 3,8%, Fe 400 частей на миллион, Mn 200 частей на миллион, Zn 200 частей на миллион, Cu 100 частей на миллион, B 100 частей на миллион и Mo 10 частей на миллион] вносили в фазу кущения. Горшки постоянно хорошо поливали и регулярно применяли жидкую мочевину (46% N). Экспериментальные единицы были организованы в полностью случайном порядке и размещены на открытом воздухе в преобладающих полевых условиях в течение обычного зимнего вегетационного периода на л.ригидум .
03% и Mn 0,08%] и жидкие удобрения [N 19% (NH 2 15%, NH 4 1,9%, NO 3 2,1%), P 8%, K 16%, Mg 1,2%, S 3,8%, Fe 400 частей на миллион, Mn 200 частей на миллион, Zn 200 частей на миллион, Cu 100 частей на миллион, B 100 частей на миллион и Mo 10 частей на миллион] вносили в фазу кущения. Горшки постоянно хорошо поливали и регулярно применяли жидкую мочевину (46% N). Экспериментальные единицы были организованы в полностью случайном порядке и размещены на открытом воздухе в преобладающих полевых условиях в течение обычного зимнего вегетационного периода на л.ригидум .
Через 2 месяца роста надземную вегетативную биомассу и площадь листьев отдельных целевых растений для каждого соответствующего генотипа ACCase RR или SS определяли, как указано выше. Оценивали надземную биомассу соседних растений пшеницы. Каждая экспериментальная обработка имела семь повторов.
Для стандартизации различий в продуктивности данные по производству биомассы и площади листьев целевых растений в присутствии соседей выражали в процентах от этого признака в отсутствие конкуренции (Goldberg and Scheiner, 2001).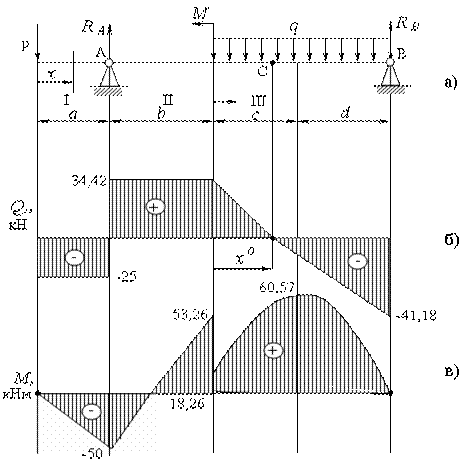 Конкурентные реакции на душу населения и на единицу размера были проанализированы с использованием гиперболической нелинейной модели для описания реакции целевых растений на увеличение плотности и биомассы соседних растений пшеницы (Weiner, 1982; Goldberg and Werner, 1983; Goldberg and Fleetwood, 1987). ):
Конкурентные реакции на душу населения и на единицу размера были проанализированы с использованием гиперболической нелинейной модели для описания реакции целевых растений на увеличение плотности и биомассы соседних растений пшеницы (Weiner, 1982; Goldberg and Werner, 1983; Goldberg and Fleetwood, 1987). ):
, где G представляет собой признак приспособленности (биомасса или площадь листа) целевого растения при плотности соседей или биомассе x , a представляет собой признак приспособленности целевого растения в отсутствие конкурентов (соседей) ( x = 0), а b наклон регрессии.Модель была подобрана с помощью регрессионного анализа методом наименьших квадратов с использованием программного обеспечения SigmaPlot (версия 12.0; Systat Software Inc.). Разница в росте целевого растения, объясняемая плотностью или биомассой соседей ( R 2 регрессионной модели), указывает на важность конкуренции за ресурсы по сравнению с другими факторами, влияющими на производительность целевого растения (Goldberg and Fleetwood, 1987).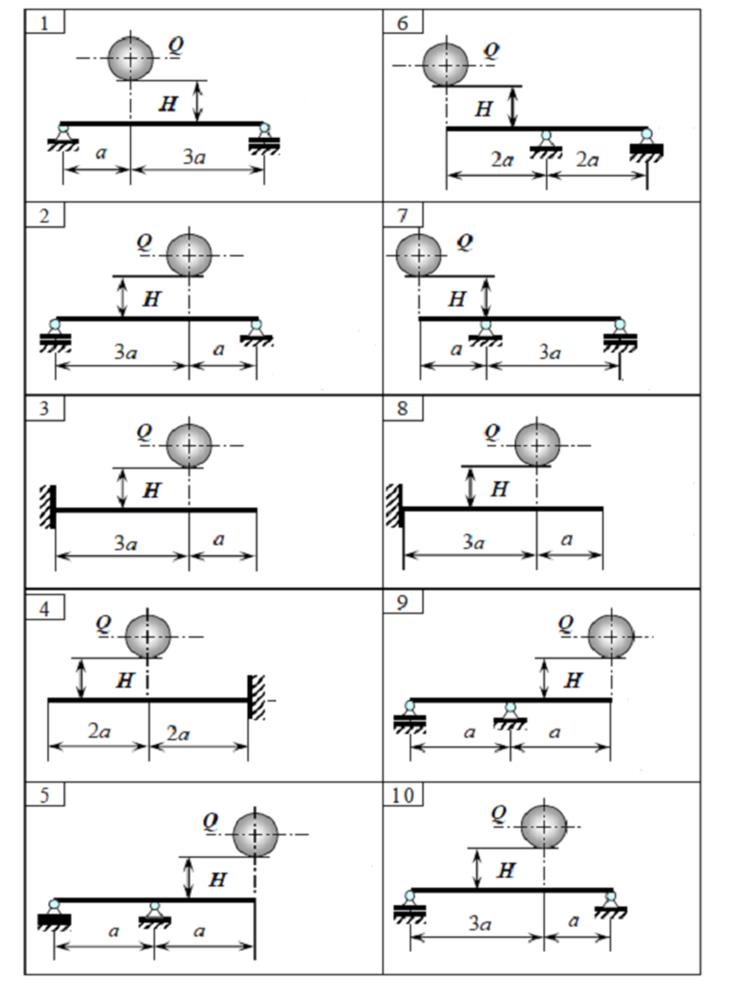
Реакция роста целевых растений (либо с устойчивостью к гербицидам АССазы, либо с аллелями дикого типа) как на увеличение количества (т.е. ответ на душу населения) и общую биомассу (т.е. на единицу размера) соседних растений пшеницы определяли после сравнения наклонов регрессии (параметр b ) с помощью однофакторного дисперсионного анализа. Более низкие и высокие наклоны обозначают сильные и слабые конкурентные реакции соответственно (Weiner, 1982; Goldberg and Werner, 1983; Goldberg, 1987). Гиперболическая модель была подобрана после логарифмического преобразования данных ( y = log [ x ]), чтобы соответствовать предположениям регрессионного анализа.
Результаты
Влияние мутаций устойчивости к гербицидам ACCase на активность и кинетику ACCase
Активность экстрагируемой АССазы была такой же в L. hardum , гомозиготные по мутации Ile-1781-Leu, как и растения дикого типа, чувствительные к гербицидам (рис. 1А). Напротив, у растений, гомозиготных по Asp-2078-Gly, была обнаружена значительно более низкая удельная активность АССазы (рис. 1А). Растения с мутацией Asp-2078-Gly показали только 70% экстрагируемой активности АССазы (специфическая активность) по сравнению с восприимчивыми растениями дикого типа. Аналогичным образом, в A. myosuroides растения, гомозиготные по мутации Asp-2078-Gly, показали значительно сниженную (40%) активность АССазы, тогда как растения с мутацией Ile-1781-Leu не показали изменений активности АССазы по сравнению с двумя чувствительный А.myosuroides популяций (рис. 1Б).
hardum , гомозиготные по мутации Ile-1781-Leu, как и растения дикого типа, чувствительные к гербицидам (рис. 1А). Напротив, у растений, гомозиготных по Asp-2078-Gly, была обнаружена значительно более низкая удельная активность АССазы (рис. 1А). Растения с мутацией Asp-2078-Gly показали только 70% экстрагируемой активности АССазы (специфическая активность) по сравнению с восприимчивыми растениями дикого типа. Аналогичным образом, в A. myosuroides растения, гомозиготные по мутации Asp-2078-Gly, показали значительно сниженную (40%) активность АССазы, тогда как растения с мутацией Ile-1781-Leu не показали изменений активности АССазы по сравнению с двумя чувствительный А.myosuroides популяций (рис. 1Б).
Рис. 1.
Активность экстрагируемой АССазы, измеренная на частично очищенных ферментных экстрактах на стадии трех-четырех листьев (A) L. hardum и (B) A. myosuroides растений гербицидов ACCase, чувствительных ( SS ), дикого типа (белая полоса) и гомозиготных устойчивых ( RR ) мутантов (Ile-1781-Leu и Asp-2078-Gly).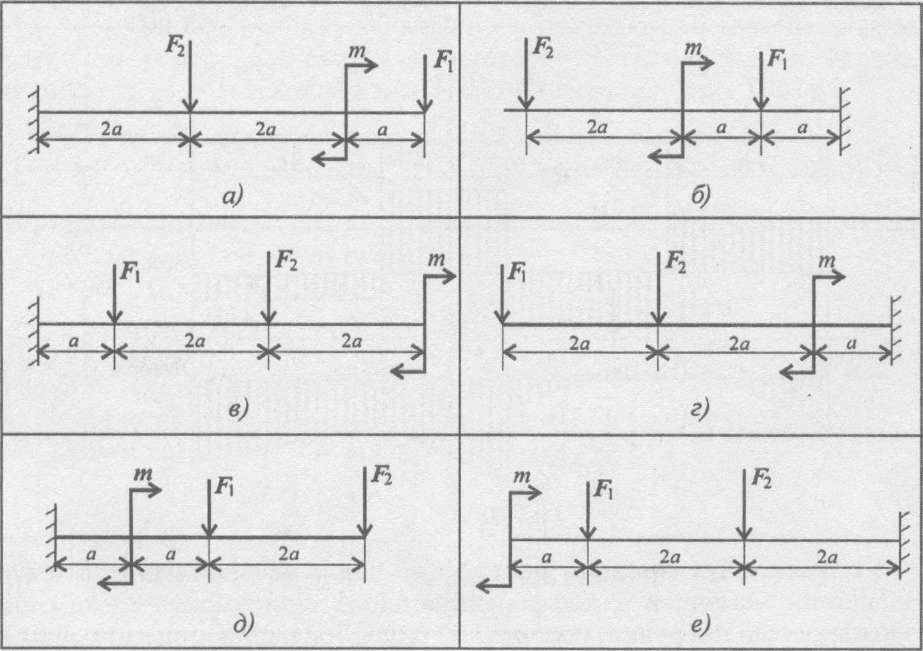 Звездочки указывают на значительные различия между средними значениями в соответствии с апостериорным тестом Даннета с использованием средних популяций, чувствительных к гербицидам ACCase, в качестве контроля (α = 0.05). НС: не имеет значения.
Звездочки указывают на значительные различия между средними значениями в соответствии с апостериорным тестом Даннета с использованием средних популяций, чувствительных к гербицидам ACCase, в качестве контроля (α = 0.05). НС: не имеет значения.
Рис. 1.
Активность экстрагируемой АССазы, измеренная на частично очищенных ферментных экстрактах стадии трех-четырех листьев (A) L. Ridum и (B) A. myosuroides растений гербицидов ACCase-чувствительных ( SS ), дикого типа (белая полоса) и гомозиготных устойчивых ( RR ) мутантов (Ile-1781-Leu и Asp-2078-Gly). Звездочки указывают на значительные различия между средними значениями в соответствии с апостериорным тестом Даннета с использованием средних популяций, чувствительных к гербицидам ACCase, в качестве контроля (α = 0.05). НС: не имеет значения.
Аффинность субстрата АССазы ( км ) к ацетил-КоА, HCO 3 — и АТФ определяли для двух мутаций резистентности и сравнивали с контролями АССазы дикого типа. Ни одна из двух мутаций ACCase не изменила значения Km , особенно для ацетил-КоА, хотя мутация Ile-1781-Leu увеличила Km (АТФ) (таблица 1). Рассчитанные Vmax под каждым субстратом также показали неизменность максимальной скорости реакции для мутантов Ile-1781-Leu, несмотря на значительное снижение значений для мутантов Asp-2078-Gly (33–48%) (табл. 1).
Ни одна из двух мутаций ACCase не изменила значения Km , особенно для ацетил-КоА, хотя мутация Ile-1781-Leu увеличила Km (АТФ) (таблица 1). Рассчитанные Vmax под каждым субстратом также показали неизменность максимальной скорости реакции для мутантов Ile-1781-Leu, несмотря на значительное снижение значений для мутантов Asp-2078-Gly (33–48%) (табл. 1).
Ошибка км (мм) и VMAX (NMOL HCO 3 — MG белок -1 мин -1 ) Значения для ацетил-COA, HCO 3 — , и АТФ-субстраты, определенные для частично очищенной АССазы из растений, гомозиготных по Ile-1781-Leu, Asp-2078-Gly и восприимчивых растений дикого типа в L. Riginum
| . | Ацетил-КоА . | ОХС 3 − . | СПС . | |||
|---|---|---|---|---|---|---|
Мутации устойчивости к ACCase .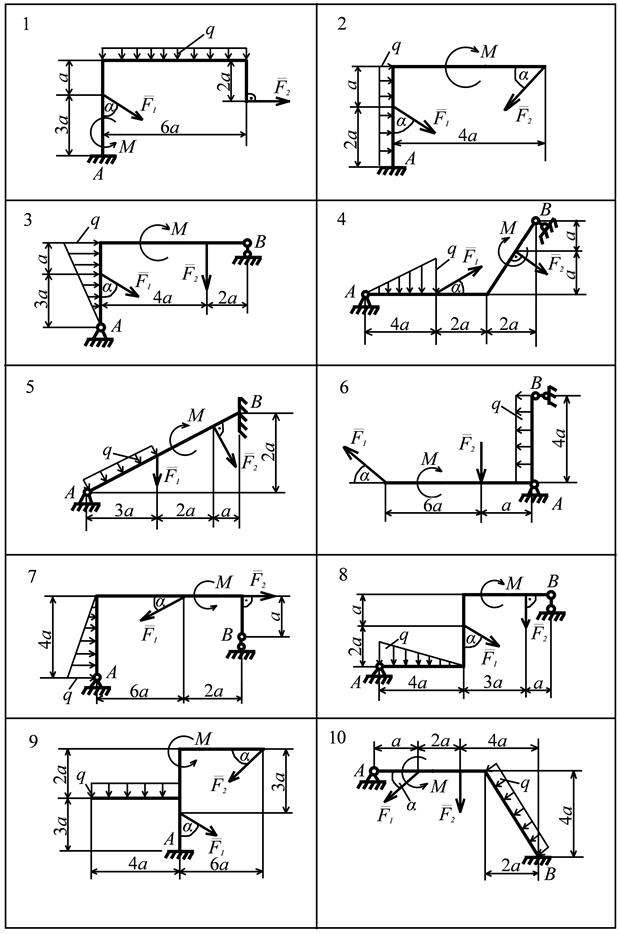 | км . | Вмакс . | км . | Вмакс . | км . | Вмакс . |
| Дикий тип | 0.091 | 35,6 | 3.5 | 38,0 | 0,116 | 38,1 |
| Иле-1 781-Лей | 0,094 | 34.7 A | 30070 ab0.158 B | A|||
| ASP-2078-GLY | 0.079 | 18,4 б | 3.3 | 25,6 б | 0,097 | 20,4 б |
. | Ацетил-КоА . | ОХС 3 − . | СПС . | |||
|---|---|---|---|---|---|---|
| Мутации устойчивости к ACCase . | км . | Вмакс . | км . | Вмакс . | км . | Вмакс . |
| дикий тип | 0.091 A | A A 38,0 A0.116 | 38,1 | |||
| Ил-тысяча семьсот восемьдесят одна-Лей | 0,094 | 34,7 | 3.5 | 33,5 AB | 0,158 B | 718.4 B B A | 25.6 B B | 0.097 | 20.4 | Очевидные км (мм) и VMAX (NMOL HCO 3 − мг белка −1 мин −1 ) значений для ацетил-КоА, HCO 3 − и субстратов АТФ, определенных для частично очищенной АЦКазы из растений, гомозиготных по Ile-1781-Leu, Asp-2078- Gly и восприимчивый дикий тип в L.твердый
 079
079 | . | Ацетил-КоА . | ОХС 3 − . | СПС
. | |||
|---|---|---|---|---|---|---|
| Мутации устойчивости к ACCase . | км . | Вмакс . | км . | Вмакс . | км . | Вмакс . |
| дикого типа | 0,091 | 35,6 | 3.5 | 38,0 | 0,116 | 38,1 |
| Иле 1781-Leu | 0,094 а | 34,7 а | 3,5 а | 33.5 ab | 0.079 | 7 18.4 B 3.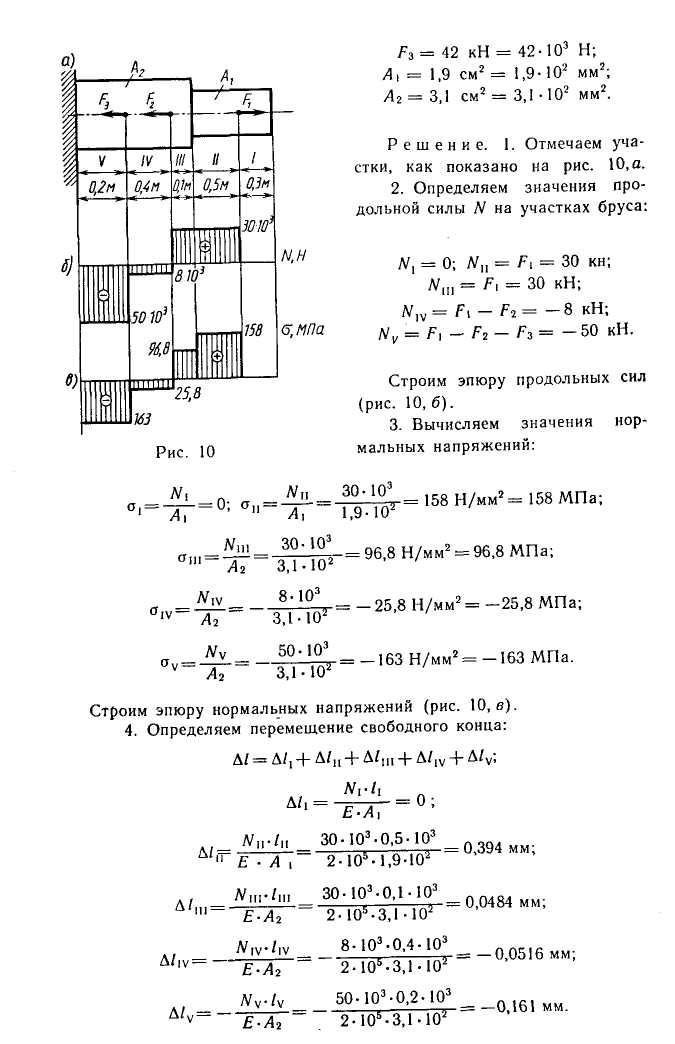 3 A 3 A | 25.6 B | 0.097 | 20.4 B |
| . | Ацетил-КоА . | ОХС 3 − . | СПС . | |||
|---|---|---|---|---|---|---|
| Мутации устойчивости к ACCase . | км . | Вмакс . | км . | Вмакс . | км . | Вмакс . |
| дикого типа | 0,091 | 35,6 | 3.5 | 38,0 | 0,116 | 38,1 |
| Иле 1781-LEU | 0. 094 A 094 A | 34.7 A 0,158 B 35.0 A | ||||
| ASP-2078-GLY | 0.079 | 9.4 B7 25.6 B | 0.097 A | 20.4 б | ||
Экспертиза на устойчивость
Выращивание изолированных растений без конкуренции
Расчетные параметры RGR, NAR и LAR не отличались среди четырех популяций, чувствительных к гербицидам ACCase ( P > 0. 05) (табл. 2). Поэтому для всех восприимчивых популяций был рассчитан средний параметр роста, который использовался в качестве эталонной оценки для дальнейших сравнений. Анализ роста показал, что растения, гомозиготные по мутации Ile-1781-Leu, демонстрировали параметры роста RGR, NAR и NAR, аналогичные чувствительным к гербицидам растениям (таблица 2). В конце вегетационного периода мутанты Ile-1781-Leu демонстрировали надземную и корневую биомассу и площадь листьев, сходные с восприимчивыми растениями дикого типа (рис. 2).
05) (табл. 2). Поэтому для всех восприимчивых популяций был рассчитан средний параметр роста, который использовался в качестве эталонной оценки для дальнейших сравнений. Анализ роста показал, что растения, гомозиготные по мутации Ile-1781-Leu, демонстрировали параметры роста RGR, NAR и NAR, аналогичные чувствительным к гербицидам растениям (таблица 2). В конце вегетационного периода мутанты Ile-1781-Leu демонстрировали надземную и корневую биомассу и площадь листьев, сходные с восприимчивыми растениями дикого типа (рис. 2).
Средние оценки RGR и компонентов NAR и LAR, связанных с генотипами L. hardum , проявляющими АССазу дикого типа, и специфическими гомозиготными ( RR ) устойчивыми мутантами (Ile-1781-Leu, Asp-2078-Gly)
| Мутация гена ACCase . | RGR (день −1 ) . | NAR (г·см −2 сут −1 ) . | LAR (см 2 г −1 )
. | ||
|---|---|---|---|---|---|
| дикий тип (S 1 ) | 0,22 (0,004) | 0,22 (0,004) | 0,0012 (3e-05) | 178 (5) | |
| Дикий Тип (S 2 ) | 0.24 ( 0.004) | 0.0013 (4E-05) | 197 (6) | ||
| дикий тип (S 3 ) | 0,24 (0,004) | 0,0013 (3E-05) | 193 (8) | ||
| Дикий тип (S 4 ) | 0,24 (0,005) | 0.0013 (5E-05) | 188 (8) | ||
| дикий тип (объединенный S 1 -S 4 ) | 0.23 A (0.004) | 0.0013 A (2e-05) | 189 (4) (4) | ||
| ILE-1781-LEU | 0. 21 A (0.005) 21 A (0.005) | 0,0012 a (4E-04) | 180 a (6) | ||
| Asp-2078-Gly | 0.16 B B (0.007) | 0.008 B (4e-04) | 190 | 190 (11) | |
| Accase Мутация гена . | RGR (день −1 ) . | NAR (г·см −2 сут −1 ) . | LAR (см 2 г −1 ) . | |
|---|---|---|---|---|
| Дикий тип (S 1 ) | 0.22 (0.004) | 0.0012 (3E-05) | 178 (5) | |
| дикий тип (S 2 ) | 0,24 (0,004) | 0,0013 (4E-05) | 197 (6) | |
| дикий тип (S 3 ) | 0,24 (0. 004) 004) | 0,0013 (3E-05) | 193 (8) | 193 (8) |
| дикий тип (S 4 ) | 0,24 (0,005) | 0,0013 (5E-05) | 188 (8) | |
| Дикий тип (объединенный S 1 –S 4 ) | 23 A (0.004) (0,004) 0,0013 (2e-05) (2e-05) A (4) 8||||
| ILE- 1781-LEU | 0.21 A (0.005) | 0.0012 A (4e-04) | 180 | (6) |
| ASP-2078-GLY | 0.16 B (0,007) | 0.008 B B (4E-04) (4E-04) | 190 (11) | (11) (11)
Средние оценки RGR и компонентов NAR и LAR, связанные с L. Gensidum Генотипы, демонстрирующие тип ACCase и специфические гомозиготные ( RR ) устойчивые мутанты (Ile-1781-Leu, Asp-2078-Gly)
Gensidum Генотипы, демонстрирующие тип ACCase и специфические гомозиготные ( RR ) устойчивые мутанты (Ile-1781-Leu, Asp-2078-Gly)
| Мутация гена ACCase . | RGR (день −1 ) . | NAR (г·см −2 сут −1 ) . | LAR (см 2 г −1 ) . | ||
|---|---|---|---|---|---|
| дикий тип (S 1 ) | 0,22 (0,004) | 0,22 (0,004) | 0,0012 (3e-05) | 178 (5) | |
| Дикий Тип (S 2 ) | 0.24 ( 0,004) | 0,0013 (4E-05) | 197 (6) | ||
| Дикий тип (S 3 ) | 0.24 (0.004) | 0.0013 (3E-05) | 193 (8) | ||
| дикий тип (S 4 ) | 0,24 (0,005) | 0,0013 (5E-05) | 188 (8) | ||
| дикий тип (объединенный S 1 -S 4 ) | 0. 23 A (0,004) 23 A (0,004) | 0,0013 A (2E-05 ) | 189 a (4) | ||
| Ile-1781-Leu | 7321 A (0,005) (0,005) | 0.0012 (4E-04) | 180 | (6) | |
| ASP-2078-GLY | 0,16 B (0,007) | 0.008 B (4e-04) | 190 (11) (11) |
| Accase Gene Mutation . | RGR (день −1 ) . | NAR (г·см −2 сут −1 ) . | LAR (см 2 г −1 ) . | ||
|---|---|---|---|---|---|
| дикий тип (S 1 ) | 0,22 (0,004) | 0,22 (0,004) | 0,0012 (3E-05) | 178 (5) | |
| Дикий Тип (S 2 ) | 0. 24 ( 0.004) 24 ( 0.004) | 0.0013 (4E-05) | 197 (6) | ||
| дикий тип (S 3 ) | 0,24 (0,004) | 0,0013 (3E-05) | 193 (8) | ||
| Дикий тип (S 4 ) | 0.24 (0,005) | 0,0013 (5E-05) | 188 (8) | ||
| дикий тип (объединенный S 1 -S 4 ) | 0,23 A ( 0,004) | 0.0013 (2e-05) | (4) (4) | ||
| ILE-1781-LEU | 0.21 A (0,005) | 0.0012 A (4E-04) (4E-04) | 180 | (6)||
| ASP-2078-GLY | 0.16 B (0,007) | (0,007)0,008 B (4E-04) | 190 a (11) |
Рис. 2.
2.
Надземная и корневая биомасса и площадь листа, продуцируемые растениями L. hardum восприимчивых диких типов ACCase ( SS ) (белая полоса) и гомозиготных устойчивых ( RR ) ( Ile-1781-Leu, Asp-2078-Gly) мутантов через 24 дня роста в отсутствие конкуренции за растительные ресурсы.Признаки роста, соответствующие растениям, несущим аллели, чувствительные к гербицидам ACCase, представляют собой средние оценки, полученные для всех восприимчивых популяций дикого типа (S 1 , S 2 , S 3 и S 4 ). Вертикальные полосы обозначают SE среднего (n = 15). Звездочки указывают на значительные различия между средними значениями в соответствии с апостериорным критерием Даннета с использованием средних популяций, чувствительных к гербицидам ACCase, в качестве контроля (α = 0,05). НС: не имеет значения.
Рис.2.
Надземная и корневая биомасса и листовая площадь, продуцируемые растениями L. Rigidum восприимчивых растений дикого типа ACCase ( SS ) (белая полоса) и гомозиготных устойчивых ( RR ) (Ile-1781) -Leu, Asp-2078-Gly) мутантов через 24 дня роста в отсутствие конкуренции за растительные ресурсы.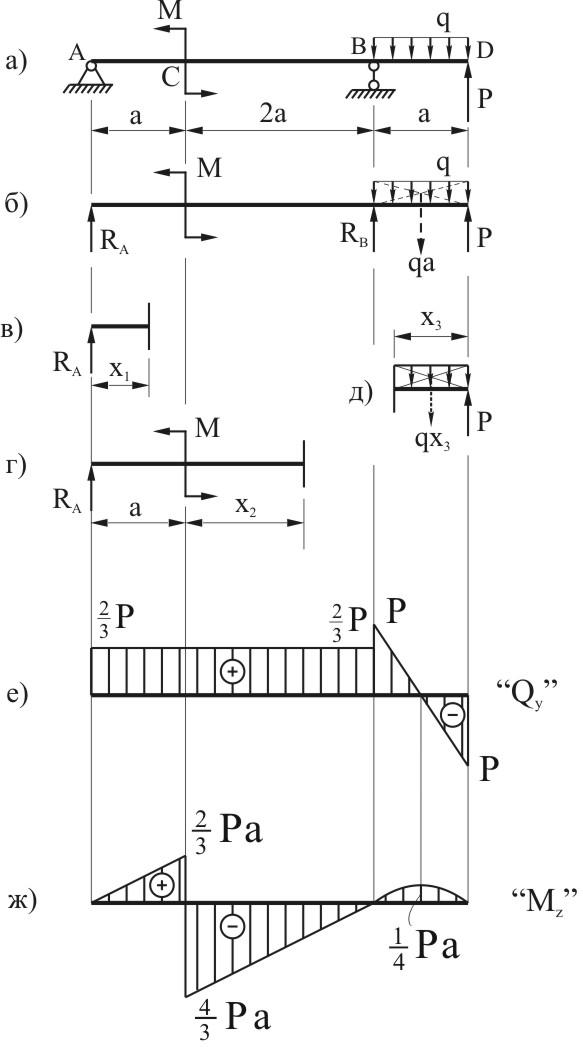 Признаки роста, соответствующие растениям, несущим аллели, чувствительные к гербицидам ACCase, представляют собой средние оценки, полученные для всех восприимчивых популяций дикого типа (S 1 , S 2 , S 3 и S 4 ).Вертикальные полосы обозначают SE среднего (n = 15). Звездочки указывают на значительные различия между средними значениями в соответствии с апостериорным критерием Даннета с использованием средних популяций, чувствительных к гербицидам ACCase, в качестве контроля (α = 0,05). НС: не имеет значения.
Признаки роста, соответствующие растениям, несущим аллели, чувствительные к гербицидам ACCase, представляют собой средние оценки, полученные для всех восприимчивых популяций дикого типа (S 1 , S 2 , S 3 и S 4 ).Вертикальные полосы обозначают SE среднего (n = 15). Звездочки указывают на значительные различия между средними значениями в соответствии с апостериорным критерием Даннета с использованием средних популяций, чувствительных к гербицидам ACCase, в качестве контроля (α = 0,05). НС: не имеет значения.
Гомозиготные мутанты Asp-2078-Gly продемонстрировали значительно сниженный RGR, обусловленный снижением NAR, но не LAR (таблица 2), что выражает более высокую стоимость устойчивости на 30% и 38%, связанную с RGR и NAR, соответственно (таблица 2) . Сниженные оценки RGR, связанные с растениями, гомозиготными по мутации Asp-2078-Gly, объясняются меньшей надземной (снижение на 74%), биомассой корней (снижение на 78%) и площадью листьев (снижение на 74%) по сравнению с восприимчивыми особями. (Инжир.2).
(Инжир.2).
Выращивание растений на условиях конкуренции
Гиперболическая модель адекватно объяснила изменения реакции роста (т.е. биомассы, площади листьев) целевых растений на увеличение плотности (реакция на душу населения) и биомассы (реакция на единицу размера) соседних растений пшеницы ( R 2 = 0,64 – 0,91, P < 0,0001). Конкурентные ответы (т.е. способность сохраняться и производить биомассу в присутствии конкурентов) растений, связанных с двумя специфическими мутациями устойчивости к АССазному гербициду, оценивали путем сравнения оценок наклона регрессии в присутствии пшеницы: чем круче наклон (более высокое значение), тем слабее конкурентная реакция.Поскольку наблюдаемые конкурентные реакции целевых растений на душу населения и на единицу размера были схожими, показаны только последние. Конкурентные реакции целевых растений L.rigidum на пшеницу варьировались в зависимости от конкретной мутации устойчивости к гербицидам ACCase, экспрессируемой растениями.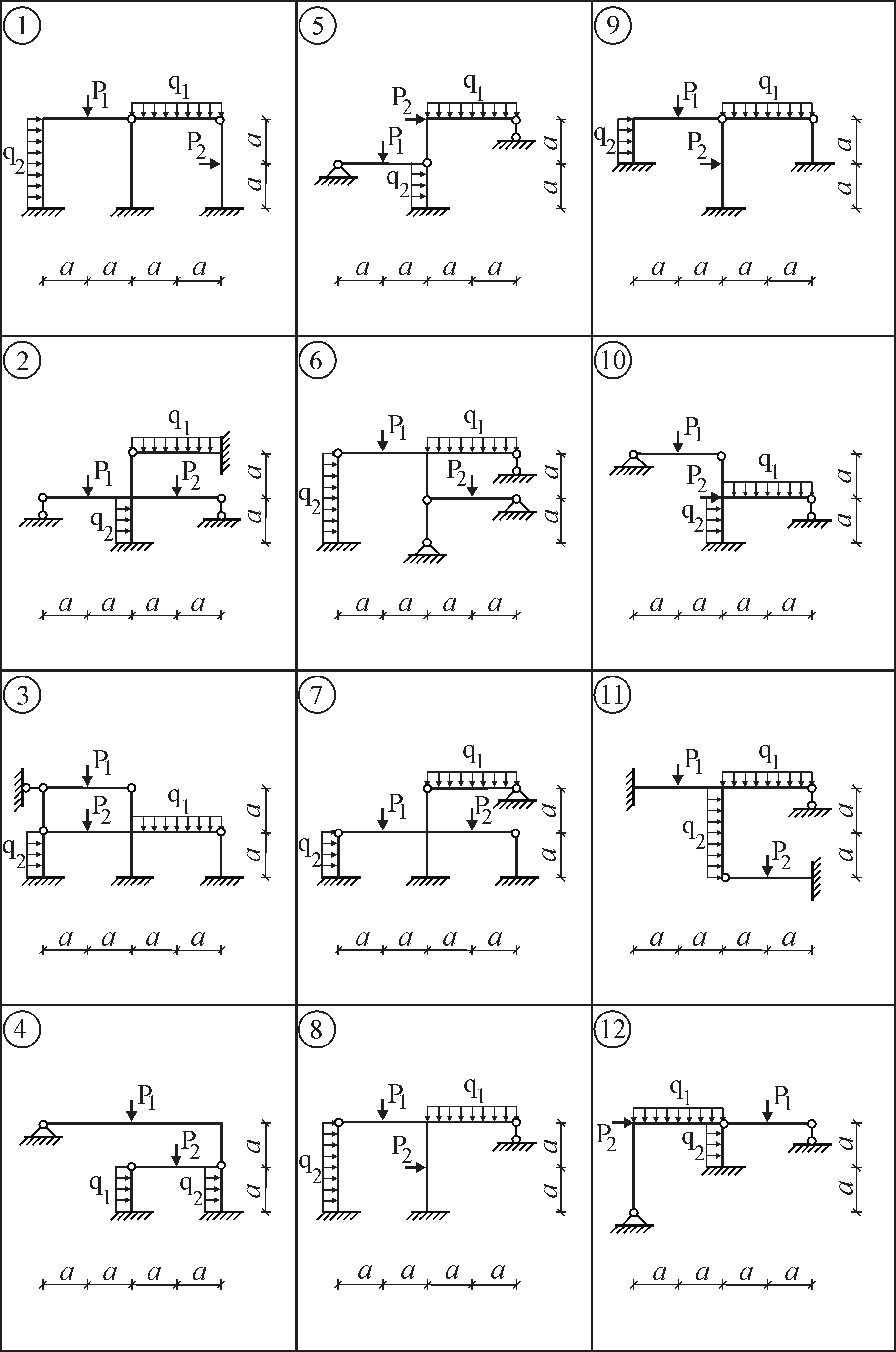
Как и ожидалось, увеличение плотности пшеницы отрицательно коррелировало с перехваченной радиацией целевыми растениями L. Ridium (рис. 3). Это привело к уменьшению размеров L.hardum с усиливающейся конкуренцией с пшеницей (рис. 4). Однако, как видно из наклона регрессий, снижение как воздушной биомассы, так и площади листьев у растений, гомозиготных по мутации Ile-1781-Leu, было сходным с таковым у растений дикого типа, чувствительных к гербицидам (рис. 4A-C). ).
Рис. 3.
Фотосинтетически активное излучение (мкмоль м −2 с −1 ), перехваченное целевыми мутантами ACCase (1781-Leu, 2078-Gly) и восприимчивыми растениями дикого типа в зависимости от увеличения густота пшеницы (0–600 растений м –2 ).Оценки проводились через 37 дней после начала эксперимента.
Рис. 3.
Фотосинтетически активное излучение (мкмоль·м −2 с −1 ), перехваченное целевыми мутантами ACCase (1781-Leu, 2078-Gly) и восприимчивыми растениями дикого типа в зависимости от повышение плотности пшеницы (0–600 растений м –2 ).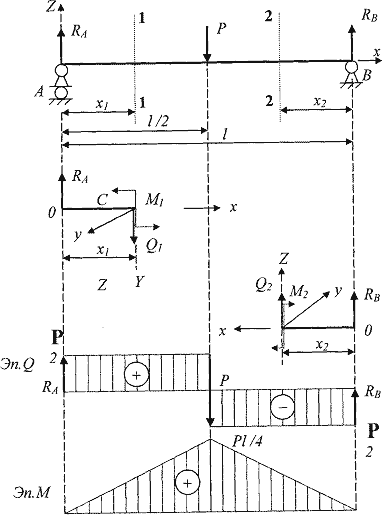 Оценки проводились через 37 дней после начала эксперимента.
Оценки проводились через 37 дней после начала эксперимента.
Рис. 4.
Вариации надземной вегетативной биомассы и площади листьев целевых устойчивых к гербицидам мутантов АССазы ( RR ) Ile-1781-Leu (●) и Asp-2078-Gly (▲) и восприимчивых диких тип ( SS ) (○) особи (S 1 , S 2 , S 3 , S 4 ) при увеличении биомассы растений-соседей пшеницы.Сравнение наклонов регрессии (параметр b ) определяет иерархию в конкурентных ответах целевых растений на единицу размера соседним растениям пшеницы.
Рис. 4.
Вариации надземной вегетативной биомассы и площади листьев целевых устойчивых к гербицидам мутантов АССазы ( RR ) Ile-1781-Leu (●) и Asp-2078-Gly (▲) и восприимчивых диких -тип ( SS ) (○) особи (S 1 , S 2 , S 3 , S 4 ) при увеличении биомассы растений-соседей пшеницы.Сравнение наклонов регрессии (параметр b ) определяет иерархию в конкурентных ответах целевых растений на единицу размера соседним растениям пшеницы.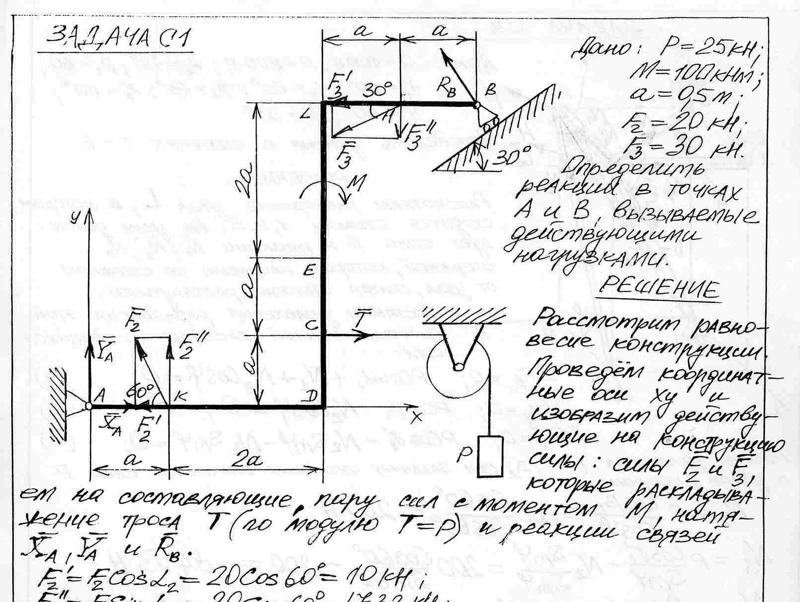
По сравнению с биомассой, полученной чувствительными к гербицидам растениями-мишенями, мутанты Asp-2078-Gly одновременно продуцировали меньшую биомассу при усилении конкуренции с пшеницей (рис. 4B-D). Это было обозначено более крутыми склонами, оцененными после регрессии. Уменьшение размера целевых растений мутации Asp-2078-Gly не сопровождалось уменьшением площади листа, как у особей, чувствительных к гербициду АССазы (рис.4Д). Дополнительным признаком слабого конкурентного ответа (т.е. надземной биомассы), связанного с мутацией устойчивости к Asp-2078-Gly, была гибель четырех целевых растений при конкуренции с урожаем на 200 и 480 растениях пшеницы м -2 .
Обсуждение
Динамика эволюции устойчивости растений к гербицидам зависит от признаков приспособленности, обусловленных мутациями устойчивости к гербицидам, как в присутствии, так и в отсутствие селекции гербицидов (Jasieniuk et al., 1996).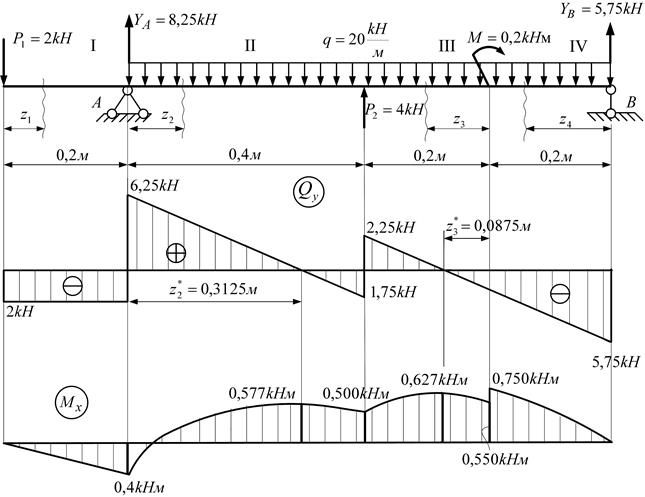 Затраты на устойчивость, связанные с аллелями устойчивости к гербицидам, учитывают вероятность фиксации новых мутаций устойчивости в популяциях сорняков, не подвергавшихся селекции гербицидами, или популяциях, в которых селекция была прекращена.
Затраты на устойчивость, связанные с аллелями устойчивости к гербицидам, учитывают вероятность фиксации новых мутаций устойчивости в популяциях сорняков, не подвергавшихся селекции гербицидами, или популяциях, в которых селекция была прекращена.
У однолетних видов, таких как L. Rigidum , изменения скорости роста и размеров растений положительно коррелируют с изменениями репродуктивных признаков (Bazzaz et al. , 1987; Weiner, 2004; Vila-Aiub et al. , 2009a). ; Weiner и др., 2009 г.). В настоящем исследовании оценивались различные признаки роста, определяющие общую приспособленность популяции, связанную с двумя специфическими мутациями гена устойчивости к АССазе к гербицидам у L.rigidum . Также была исследована корреляция между влиянием этих мутаций на активность и кинетику ACCase и этими компонентами приспособленности. Были проведены сравнения между гомозиготными растениями с мутациями ACCase Ile-1781-Leu или Asp-2078-Gly и восприимчивыми растениями дикого типа из четырех разных L.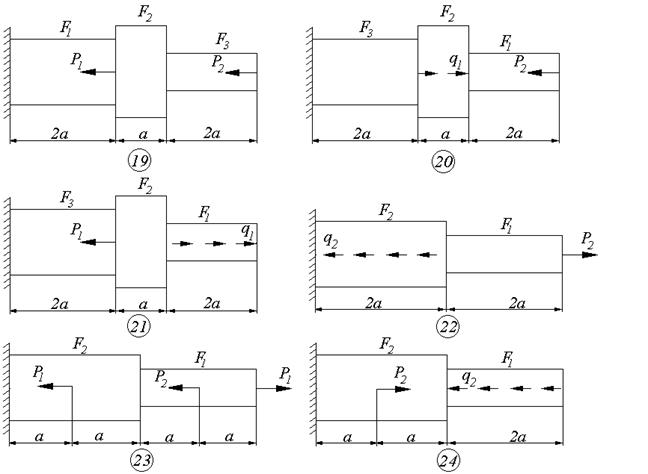 ригидум популяций.
ригидум популяций.
Экспериментальные результаты показывают, что (i) мутации устойчивости к Ile-1781-Leu и Asp-2078-Gly могут обеспечивать устойчивость к гербицидам ACCase без значительного вмешательства в нормальное связывание субстрата ( Km ), (ii) уровни АССазная активность и каталитическая способность ( Vmax ), связанные с каждой мутацией, могут быть коррелированы с величиной затрат на устойчивость, и (iii) затраты на устойчивость связаны с мутацией Asp-2078-Gly, но не с мутацией Ile-1781-Leu. .
ACCase Мутация Ile-1781-Leu: не влияет на кинетику ACCase и не требует затрат на резистентность Гомозиготные резистентные мутанты Ile-1781-Leu имели схожие реакции RGR-NAR и конкуренции за ресурсы по сравнению с чувствительными к гербицидам растениями, обладающими АССазой дикого типа. Эти результаты согласуются с двумя предыдущими исследованиями, показывающими незначительные затраты на приспособляемость, связанные с мутацией Ile-1781-Leu у L. hardum и A. myosuroides (Vila-Aiub et al., 2005 г.; Менчари и др. , 2008 г.). При интрогрессии в Setaria italica мутанты Ile-1781-Leu даже показали преимущество в приспособленности в отсутствие селекции гербицидов (Wang et al. , 2010). Вполне вероятно, что отсутствие стоимости резистентности, связанной с этой мутацией Ile-1781-Leu в L. Rigidum , связано с тем, что эта мутация не оказывает неблагоприятного воздействия на кинетику фермента АССазы. Сродство к субстрату Ile-1781-Leu ( Km ) и скорость образования продукта ( Vmax ) были аналогичны восприимчивой АССазе дикого типа (Vila-Aiub et al., 2009b; Ю и др. , 2010). В растениях A. myosuroides также не было выявлено неблагоприятного воздействия мутации Ile-1781-Leu на активность АССазы (фиг. 1В).
hardum и A. myosuroides (Vila-Aiub et al., 2005 г.; Менчари и др. , 2008 г.). При интрогрессии в Setaria italica мутанты Ile-1781-Leu даже показали преимущество в приспособленности в отсутствие селекции гербицидов (Wang et al. , 2010). Вполне вероятно, что отсутствие стоимости резистентности, связанной с этой мутацией Ile-1781-Leu в L. Rigidum , связано с тем, что эта мутация не оказывает неблагоприятного воздействия на кинетику фермента АССазы. Сродство к субстрату Ile-1781-Leu ( Km ) и скорость образования продукта ( Vmax ) были аналогичны восприимчивой АССазе дикого типа (Vila-Aiub et al., 2009b; Ю и др. , 2010). В растениях A. myosuroides также не было выявлено неблагоприятного воздействия мутации Ile-1781-Leu на активность АССазы (фиг. 1В).
Растения, гомозиготные по мутации Asp-2078-Gly, демонстрировали сниженные параметры роста RGR-NAR, что подразумевает отрицательную связь с эффективностью растения в улавливании света, ассимиляции CO 2 и/или хранении фотоассимилятов. У особей, обладающих мутацией Asp-2078-Gly, было обнаружено значительное уменьшение площади листьев, надземной и корневой биомассы. В условиях конкуренции растения с мутацией Asp-2078-Gly также демонстрировали нарушение роста в условиях конкуренции за ресурсы, проявляющееся в виде слабого конкурентного ответа на пшеницу по сравнению с растениями с восприимчивой АССазой дикого типа. Слабые конкурентные ответы растений с мутацией Asp-2078-Gly согласовывались с увеличением как плотности пшеницы, так и биомассы. Этот результат указывает на наличие признаков растений, связанных с мутацией Asp-2078-Gly, отличных от «уменьшенного размера растения», которые способствуют слабой конкурентной реакции, поскольку различия в конкурентных реакциях, возникающие только из-за разных размеров растений, не должны быть очевидными при корректировке с помощью размер (соседний вес) (Goldberg and Scheiner, 2001).В конкурентных условиях, когда свет становится самым ограничивающим ресурсом [т.е. только 6–13% доступной фотосинтетически активной радиации наблюдалось при высокой плотности пшеницы (200–480 растений м -2 )], ожидается, что растения с пониженной NAR будут неэффективно использовать радиацию.
У особей, обладающих мутацией Asp-2078-Gly, было обнаружено значительное уменьшение площади листьев, надземной и корневой биомассы. В условиях конкуренции растения с мутацией Asp-2078-Gly также демонстрировали нарушение роста в условиях конкуренции за ресурсы, проявляющееся в виде слабого конкурентного ответа на пшеницу по сравнению с растениями с восприимчивой АССазой дикого типа. Слабые конкурентные ответы растений с мутацией Asp-2078-Gly согласовывались с увеличением как плотности пшеницы, так и биомассы. Этот результат указывает на наличие признаков растений, связанных с мутацией Asp-2078-Gly, отличных от «уменьшенного размера растения», которые способствуют слабой конкурентной реакции, поскольку различия в конкурентных реакциях, возникающие только из-за разных размеров растений, не должны быть очевидными при корректировке с помощью размер (соседний вес) (Goldberg and Scheiner, 2001).В конкурентных условиях, когда свет становится самым ограничивающим ресурсом [т.е. только 6–13% доступной фотосинтетически активной радиации наблюдалось при высокой плотности пшеницы (200–480 растений м -2 )], ожидается, что растения с пониженной NAR будут неэффективно использовать радиацию.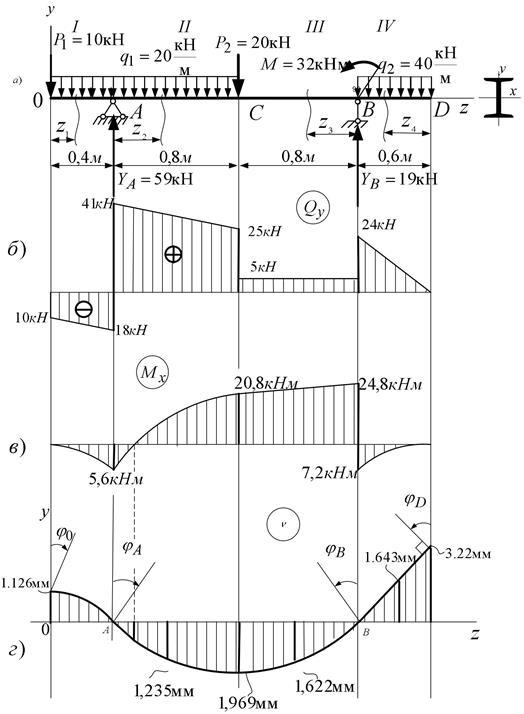
Учитывая, что репродуктивная способность растений является функцией RGR и количества ресурсов, выделяемых на вегетативную биомассу, наблюдаемое уменьшение размера, связанное с мутацией устойчивости к Asp-2078-Gly у растений, выращенных без конкуренции и с конкуренцией, вероятно, приведет к сокращению по плодовитости по сравнению с особями с АССазой дикого типа (Bazzaz et al., 1987; Вайнер, 2004 г.; Вайнер и др. , 2009 г.). Это особенно верно для однолетних видов, таких как L. Rigidum , у которых уменьшенные размеры растений и RGR положительно коррелируют со сниженными репродуктивными признаками (Vila-Aiub et al. , 2009a).
Ясно, что мутация устойчивости Asp-2078-Gly делает ACCase устойчивой к гербицидам ACCase. Эта мутация не изменяет сродство к субстрату АЦКазы ( Km ), но наблюдается почти 50% снижение скорости АЦКазы ( Vmax ) для катализа образования малонил-КоА за счет природных субстратов (ацетил-КоА, АТФ). и HCO 3 ).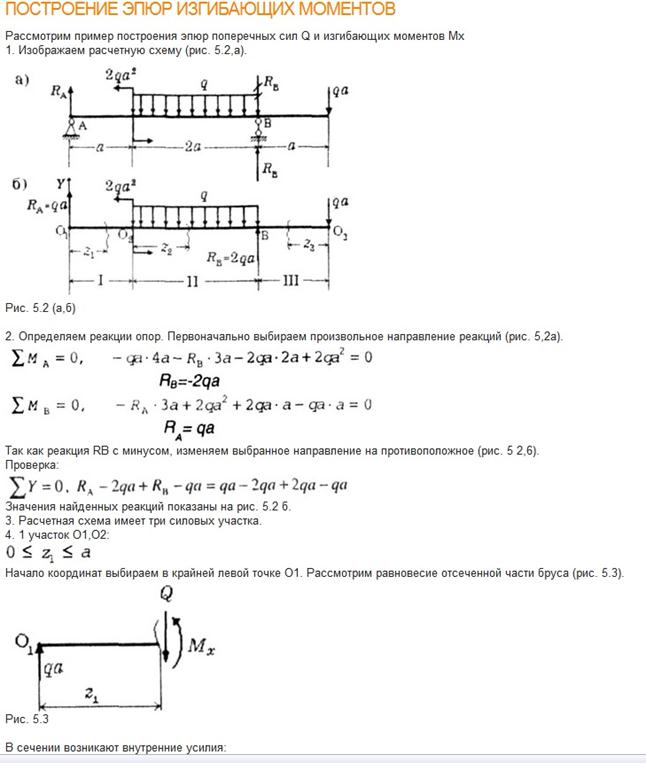 Общее снижение специфической активности АССазы (30%) и Vmax может привести к нехватке липидов, доступных для быстрого роста, и коррелировать с нарушением реакции роста, наблюдаемым у растений, гомозиготных по мутации ACCase Asp-2078-Gly.
Общее снижение специфической активности АССазы (30%) и Vmax может привести к нехватке липидов, доступных для быстрого роста, и коррелировать с нарушением реакции роста, наблюдаемым у растений, гомозиготных по мутации ACCase Asp-2078-Gly.
Мутация Asp-2078-Gly в A. myosuroides также связана с затратами на резистентность (Menchari et al. , 2008). Как показано здесь (Fig. 1B), правдоподобным объяснением этой сниженной приспособленности является нарушение активности ACCase, связанное с этой конкретной мутацией сайта-мишени.
Взгляд на эволюцию устойчивости к гербицидам ACCase
Условия, благоприятствующие быстрой эволюции и закреплению новых аллелей устойчивости к гербицидам на целевом участке в популяциях сорняков, включают как незначительные затраты на устойчивость ( RC , см. уравнение 1), проявляющиеся при смягчении отбора гербицидов, так и значительное преимущество в приспособленности или защиту растений при отборе гербицидов (Вила-Аюб и др. , 2009b). Предыдущие исследования характеризовали высокий уровень устойчивости к некоторым арилоксифеноксипропионатным и циклогександионовым гербицидам ACCase, обусловленным мутациями Ile-1781-Leu и Asp-2078-Gly у различных видов (Délye et al., 2005 г.; Делье и др. , 2008 г.; Ю и др. , 2007b).
, 2009b). Предыдущие исследования характеризовали высокий уровень устойчивости к некоторым арилоксифеноксипропионатным и циклогександионовым гербицидам ACCase, обусловленным мутациями Ile-1781-Leu и Asp-2078-Gly у различных видов (Délye et al., 2005 г.; Делье и др. , 2008 г.; Ю и др. , 2007b).
В отсутствие селекции гербицидов отсутствие затрат на устойчивость предполагает, что не существует очевидных ограничений для сохранения и закрепления аллеля устойчивости ACCase Ile-1781-Leu в популяциях. Эмпирические данные показывают, что аллель устойчивости Ile-1781-Leu был зафиксирован у некоторых видов трав и обнаружен у древних A. myosuroides особей, никогда не подвергавшихся воздействию гербицидов (Délye and Michel, 2005; Délye et al., 2013). Напротив, прогнозируется, что окружающая среда создаст больше ограничений для устойчивости аллеля ACCase Asp-2078-Gly после ослабления отбора гербицидов. Таким образом, при гербицидной селекции ожидается, что мутации Ile-1781-Leu и Asp-2078-Gly увеличат свою популяционную частоту. После прекращения отбора мутация Ile-1781-Leu с большей вероятностью сохранится, чем мутация Asp-2078-Gly.
После прекращения отбора мутация Ile-1781-Leu с большей вероятностью сохранится, чем мутация Asp-2078-Gly.
Благодарности
AHRI финансируется Австралийской корпорацией исследований и развития зерна (GRDC).MMVA была поддержана стипендией Endeavour (Департамент образования, правительство Австралии). Мы благодарим доктора Кристофа Делье за любезно предоставленный растительный материал A. myosuroides .
Каталожные номера
Базза
ФА
Кьяриелло
рупийКоли
ПД
Пителька
ЛФ
.1987
.Выделение ресурсов для воспроизводства и защиты
.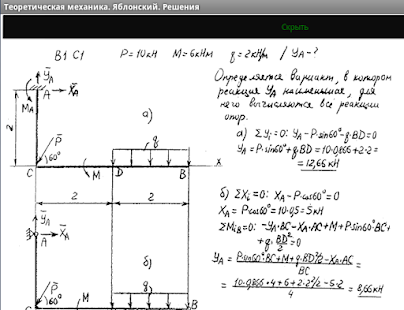
Биология
37
,58
–67
.Брэдфорд
ММ
.1976
.Быстрый и чувствительный метод количественного определения белка в микрограммах, использующий принцип связывания белка с красителем
.Аналитическая биохимия
72
,248
–254
.Каустон
ДР
Венера
ДЖК
.1981
.Биометрия роста растений
.Лондон, Великобритания
:Эдвард Арнольд
.Казенс
РД
Джилл
ГС
Спайерс
ЭДЖ
.
1997
.Комментарий: Количество выборочных популяций, необходимых для определения влияния устойчивости к гербицидам на рост и приспособленность растений
.Исследование сорняков
37
,1
–4
.Делье
С
.2005
.Устойчивость сорняков к ингибиторам ацетилкоэнзима А-карбоксилазы: обновление
.Наука о сорняках
53
,728
–746
.Делье
С
Деулвот
С
Шовель
Б
.2013
.Анализ ДНК гербарных образцов злакового сорняка Alopecurus myosuroides выявил устойчивость к гербицидам, предшествующим гербицидам
.
ПЛОС ОДИН
8
.Делье
С
Матейчек
А
Мишель
С
.2008
.Характер перекрестной устойчивости к гербицидам, ингибирующим АССазу, обусловленный мутантными изоформами АССазы у Alopecurus myosuroides Huds.(чернотравный), переобследован при рекомендованной полевой норме гербицида
.Наука о борьбе с вредителями
64
,1179
–1186
.Делье
С
Мишель
С
.2005
.«Универсальные» праймеры для ПЦР-секвенирования доменов хлоропластной ацетил-КоА-карбоксилазы трав, вовлеченных в устойчивость к гербицидам
.
Исследование сорняков
45
,323
–330
.Делье
С
Чжан
XQ
Шалопин
С
Мишель
С
Поулз
СБ
.2003
.Остаток изолейцина в карбокситрансферазном домене многодоменной ацетил-кофермента А-карбоксилазы является основным фактором, определяющим чувствительность к арилоксифеноксипропионату, но не к ингибиторам циклогександиона
.Физиология растений
132
,1716
–1723
.Делье
С
Чжан
XQ
Мишель
С
Матейчек
А
Поулз
СБ
.
2005
.Молекулярные основы чувствительности к ингибиторам ацетил-кофермента А-карбоксилазы у черной травы
.Физиология растений
137
,794
–806
.Гибсон
ДЖ
Коннолли
Дж
Хартнетт
DC
Вайденхамер
ДД
.1999
.Схемы для изучения взаимодействий между растениями в теплицах
.Журнал экологии
87
,1
–16
.Голдберг
ДЭ
.1987
.Соревнования по соседству в старопольском растительном сообществе
.Экология
68
,1211
–1223
.
Голдберг
ДЭ
.1990
.Компоненты конкуренции за ресурсы в растительных сообществах
. В:Грейс
JB
Тилман
D
, ред.Перспективы в конкурсе растений
.Сан-Диего
:Academic Press
,27
–49
.Голдберг
ДЭ
Флитвуд
л
.1987
.Конкурентный эффект и реакция четырех однолетних растений
.Журнал экологии
75
,1131
–1143
.Голдберг
ДЭ
Шайнер
СМ
.2001
. ANOVA и ANCOVA.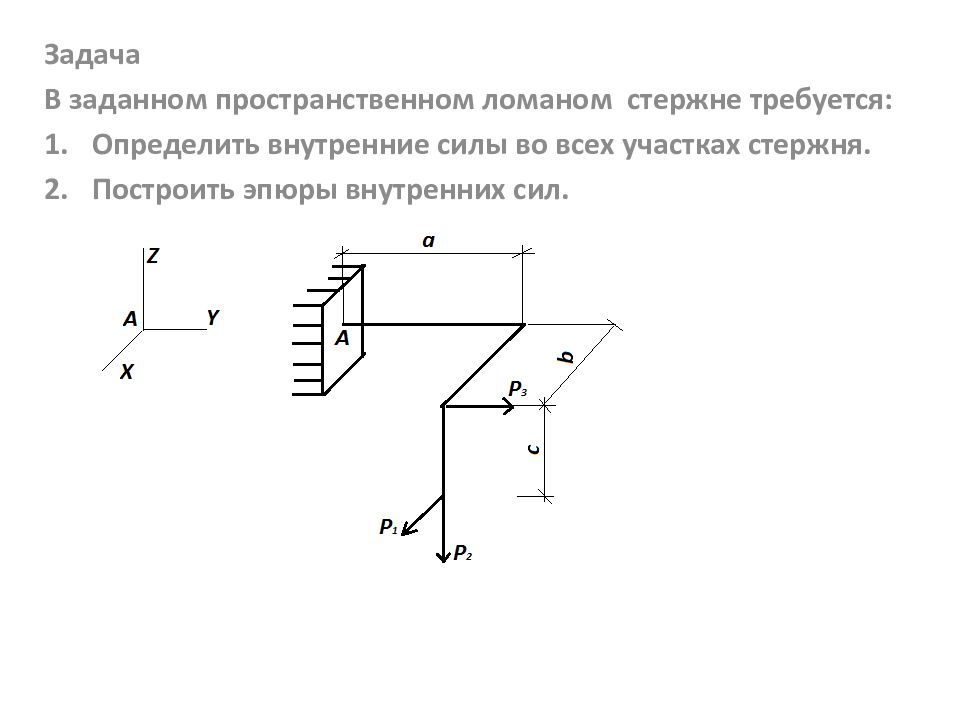 Полевые конкурсные эксперименты
Полевые конкурсные эксперименты
Scheiner
SM
Гуревич
J
, ред.Планирование и анализ экологических экспериментов
.Нью-Йорк
:Oxford University Press
,77
–98
.Голдберг
ДЭ
Вернер
ПА
.1983
.Эквивалентность конкурентов в растительных сообществах: нулевая гипотеза и полевой экспериментальный подход
.Американский журнал ботаники
70
,1098
–1104
.Грязь
JP
Охота
Р
.1975
.Относительная скорость роста: ее ареал и адаптивное значение в местной флоре
.Экологический журнал
63
,393
–422
.
Хоффманн
ВА
Путер
Х
.2002
.Избежание систематической ошибки при расчетах относительного темпа роста
.Анналы ботаники
90
,37
–42
.Охота
Р
.1982
.Кривые роста растений. Функциональный подход к анализу роста растений
.Лондон, Великобритания
:Edward Arnold Ltd
.Охота
Р
Каустон
ДР
Шипли
Б
Аскью
АП
.
2002
.Современный инструмент для классического анализа роста растений
.Анналы ботаники
90
,485
–488
.Ясенюк
М
Брюле-Бабель
АЛ
Моррисон
В
.1996
.Эволюция и генетика устойчивости сорняков к гербицидам
.Наука о сорняках
44
,176
–193
.Каундун
нержавеющая сталь
.2014
.Устойчивость к гербицидам, ингибирующим ацетил-КоА-карбоксилазу
.Наука о борьбе с вредителями
70
,1405
–1417
.Кешткар
Е
Матиассен
СК
Мох
СР
Кудск
Р
.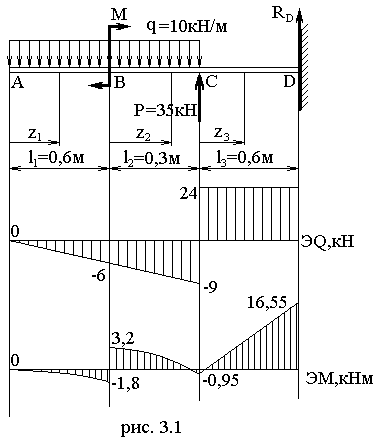
2015
.Профиль устойчивости популяций, устойчивых к гербицидам Alopecurus myosuroides (чернотравье) в Дании
.Средства защиты растений
69
,83
–89
.Мэлоун
ДжМ
Бутсалис
Р
Бейкер
Дж
Престон
С
.2013
.Распределение устойчивых к гербицидам аллелей ацетилкоэнзима карбоксилазы Lolium Rigidum в посевных площадях Южной Австралии
.Исследование сорняков
54
,78
–86
.Менчари
Д
Шовель
Б
Дарменси
Х
Делье
С
.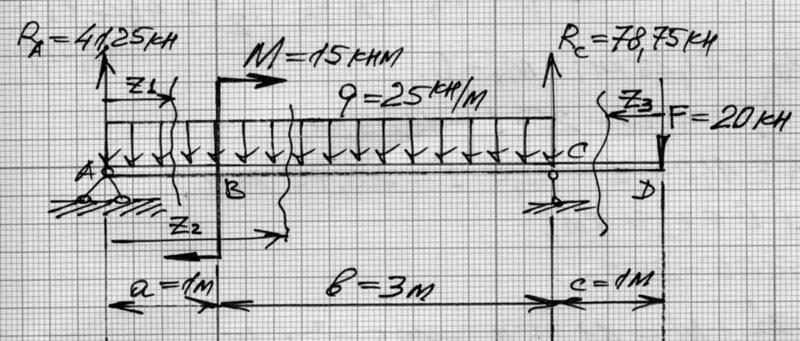
2008
.Затраты на приспособляемость, связанные с тремя мутантными аллелями ацетил-коэнзима А-карбоксилазы, обеспечивающими устойчивость к гербицидам у черной травы Alopecurus myosuroides
.Журнал прикладной экологии
45
,939
–947
.Оуэн
МДж
Уолш
МДж
Ллевелин
РС
Поулз
СБ
.2007
.Широкое распространение множественной устойчивости к гербицидам в популяциях однолетнего райграса Западной Австралии ( Lolium Rigidum )
.Австралийский журнал сельскохозяйственных исследований
58
,711
–718
.Путер
Х
Нагель
О
.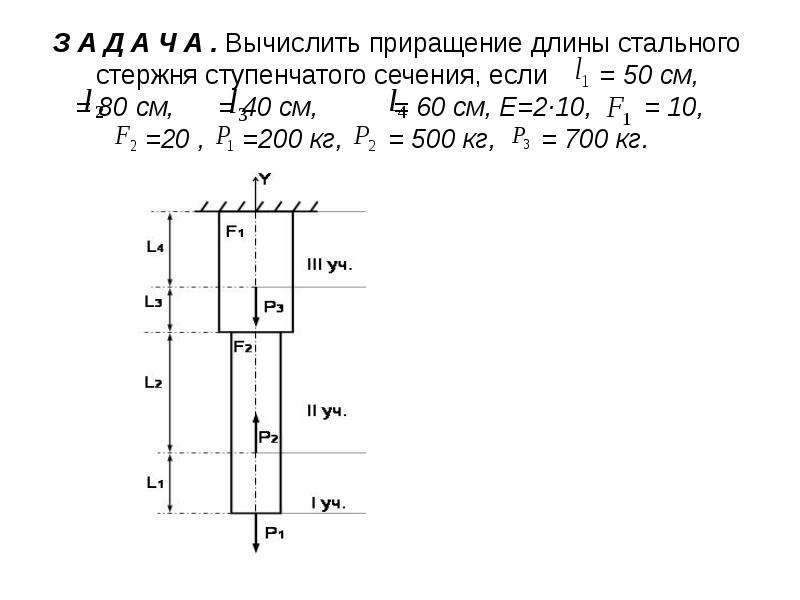
2000
.Роль распределения биомассы в ростовой реакции растений на различные уровни света, CO2, питательных веществ и воды: количественный обзор
.Австралийский журнал физиологии растений
27
,595
–607
.Поулз
СБ
Ю
В
.2010
.Эволюция в действии: растения, устойчивые к гербицидам
.Ежегодный обзор биологии растений
61
,317
–347
.Пуррингтон
КБ
Бергельсон
Дж
.1999
.Изучение физиологических основ затрат на устойчивость к гербицидам у Arabidopsis thaliana
.Американский натуралист
154
,S82
–S91
.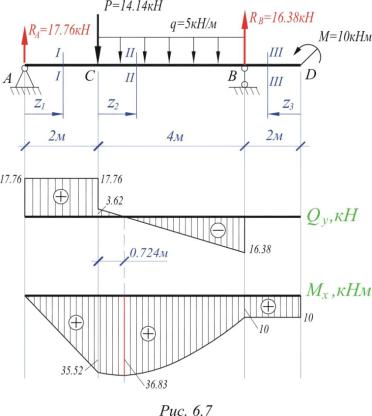
Ру
Ф
Гаскес
Дж
Ребауд
х
.2004
.Преобладание устойчивости к гербицидам в нескольких мутантных линиях Arabidopsis thaliana
.Генетика
166
,449
–460
.Штраус
СЫ
Рули
ДЖА
Лау
ДЖА
Ирвин
РД
.2002
.Прямые и экологические издержки устойчивости к травоядным
.Тенденции в экологии и эволюции
17
,278
–285
.
Вила-Аюб
ММ
Гундель
ПЭ
Престон
С
.2015
.Экспериментальные методы оценки стоимости приспособленности растений, связанной с генами устойчивости к гербицидам
.Наука о сорняках
63
,203
–216
.Вила-Аюб
ММ
Неве
Р
Поулз
СБ
.2005
.Стоимость резистентности механизма метаболизма гербицида цитохрома P450, но не мутации сайта-мишени АССазы в множественной резистентности популяции Lolium Rigidum
.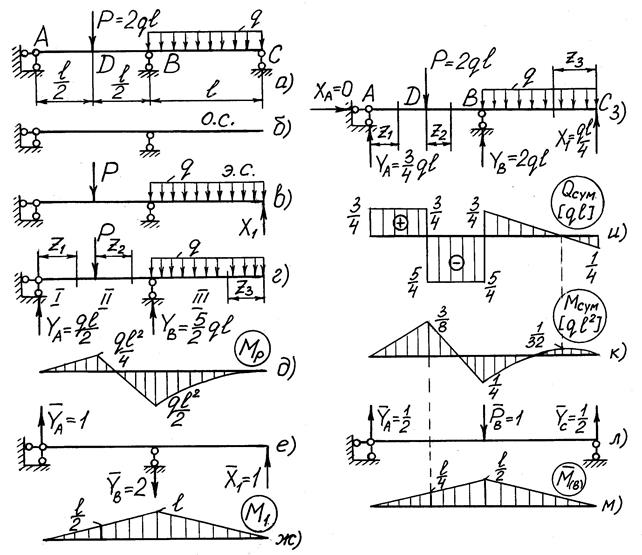
Новый фитолог
167
,787
–796
.Вила-Аюб
ММ
Неве
Р
Поулз
СБ
.2009а
.Доказательства экологических издержек усиленного метаболизма гербицидов в Lolium Rigidum
.Журнал экологии
97
,772
–780
.Вила-Аюб
ММ
Неве
Р
Поулз
СБ
.2009b
.Затраты на приспособление, связанные с развившимися аллелями устойчивости растений к гербицидам
.Новый фитолог
184
,751
–767
.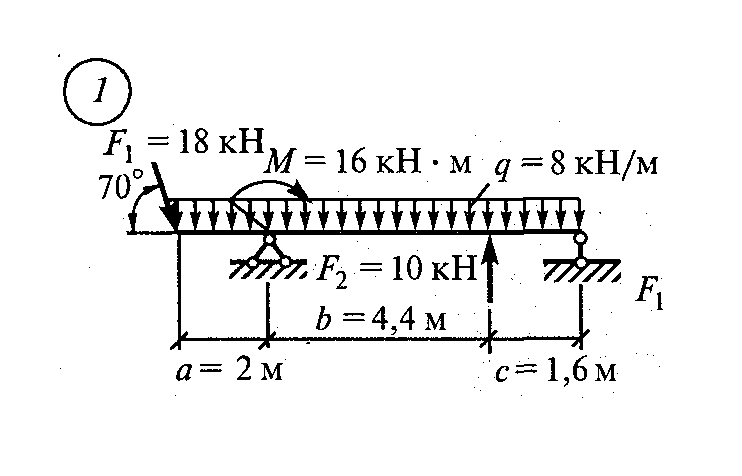
Вила-Аюб
ММ
Неве
Р
Ру
Ф
.2011
.Единый подход к оценке и интерпретации затрат на устойчивость растений
Наследственность
107
,386
–394
.Ван
Т
Пикард
ДЖК
Тиан
х
Дарменси
Х
.2010
.Устойчивый к гербицидам мутант ACCase 1781 Setaria демонстрирует более высокую приспособленность, чем дикий тип
.Наследственность
105
,394
–400
.
Вайнер
Дж
.1982
.Окрестностная модель интерференции однолетних растений
.Экология
63
,1237
–1241
.Вайнер
Дж
.2004
.Аллокация, пластичность и аллометрия у растений
.Перспективы эволюции и систематики экологии растений
6
,207
–215
.Вайнер
Дж
Кэмпбелл
ЛГ
Пино
Дж
Экарта
л
.2009
.Аллометрия размножения в популяциях растений
.
Журнал экологии
97
,1220
–1233
.Ю
В
Абдаллах
я
Хан
Х
Оуэн
М
Поулз
С
.2009
.Отдельные нецелевые механизмы обеспечивают устойчивость к глифосату, ACCase и ALS-ингибирующим гербицидам у множественно устойчивых к гербицидам Lolium Rigidum
.Планта
230
,713
–723
.Ю
В
Кэрнс
А
Поулз
С
.
2007а
.Множественная устойчивость к гербицидам глифосату, параквату и ACCase развилась у биотипа
Lolium Rigidum .Планта
225
,499
–513
.Ю
В
Коллаво
А
Чжэн
МК
Оуэн
М
Сатин
М
Поулз
СБ
.2007b
.Разнообразие мутаций ацетилкоэнзима карбоксилазы в резистентных популяциях Lolium : оценка с использованием клетодима
.Физиология растений
145
,547
–558
.Ю
В
Фризен
СО
Чжан
XQ
Поулз
СБ
.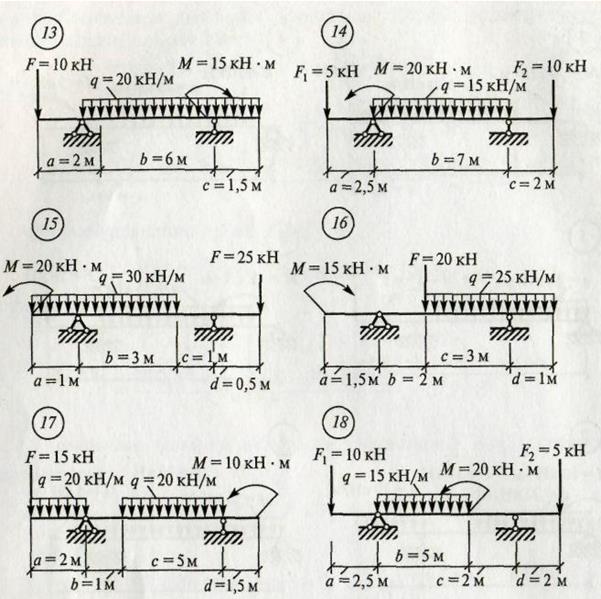
2004
.Толерантность к гербицидам, ингибирующим ацетолактатсинтазу и ацетилкоэнзим А-карбоксилазу, в Vulpia bromoides обеспечивается двумя сосуществующими механизмами устойчивости
.Биохимия и физиология пестицидов
78
,21
–30
.Ю
В
Хан
Х
Вила-Аюб
ММ
Поулз
СБ
.2010
.Устойчивость к гербицидам AHAS, вызывающая мутации: влияние на функциональность AHAS и рост растений
.Журнал экспериментальной ботаники
61
,3925
–3934
.Чжан
Х
Твил
Б
Тонг
л
.
2004
.Молекулярная основа ингибирования карбоксилтрансферазного домена ацетил-кофермент-А-карбоксилазы галоксифопом и диклофопом
.Труды Национальной академии наук США
101
,5910
–5915
.© The Author, 2015. Опубликовано Oxford University Press от имени Общества экспериментальной биологии.
Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License (http://creativecommons.org/licenses/by/3.0/), что разрешает неограниченное повторное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы.Поддержание экспоненциального роста, проводимости раствора и pH раствора в культуре раствора с низкой ионной силой с использованием управляемой компьютером системы доставки питательных веществ | Журнал экспериментальной ботаники
Аннотация
Исследования потребностей растений в питательных веществах в культуре в растворе часто использовали концентрации питательных веществ, во много раз превышающие уровни, обнаруженные в плодородных почвах, создавая искусственную среду для укоренения, которая может изменить характер усвоения питательных веществ. Метод относительной скорости добавления (RAR) решает эту проблему, обеспечивая питательные вещества в экспоненциально увеличивающихся количествах корням растений в культуре раствора. Управляемая компьютером система доставки питательных веществ RAR была разработана для снижения рабочей нагрузки и облегчения более частого добавления питательных веществ (4 раза в день), чем это возможно при добавлении вручную. В первоначальных экспериментах для здорового роста Triticum aestivum требовался минимальный фоновый раствор, содержащий 500 мкМ азота и все другие необходимые питательные вещества в оптимальных пропорциях.Это требование было снижено до 50 мкМ азота, когда содержание кальция в фоновых растворах было увеличено до 400 мкМ. Варьируя содержание аммония и нитрата как в фоновом растворе, так и в растворе для доставки, можно было контролировать вызванные растениями изменения рН в ростовых растворах. В оптимизированных растворах относительная скорость роста растений ( RGR ) порядка 0,2 г г -1 растений в день -1 поддерживалась в течение 22-дневного экспериментального периода.
Метод относительной скорости добавления (RAR) решает эту проблему, обеспечивая питательные вещества в экспоненциально увеличивающихся количествах корням растений в культуре раствора. Управляемая компьютером система доставки питательных веществ RAR была разработана для снижения рабочей нагрузки и облегчения более частого добавления питательных веществ (4 раза в день), чем это возможно при добавлении вручную. В первоначальных экспериментах для здорового роста Triticum aestivum требовался минимальный фоновый раствор, содержащий 500 мкМ азота и все другие необходимые питательные вещества в оптимальных пропорциях.Это требование было снижено до 50 мкМ азота, когда содержание кальция в фоновых растворах было увеличено до 400 мкМ. Варьируя содержание аммония и нитрата как в фоновом растворе, так и в растворе для доставки, можно было контролировать вызванные растениями изменения рН в ростовых растворах. В оптимизированных растворах относительная скорость роста растений ( RGR ) порядка 0,2 г г -1 растений в день -1 поддерживалась в течение 22-дневного экспериментального периода. Варьирование RAR обеспечило возможность выращивания растений с различными RGR s при относительно постоянных условиях электропроводности раствора и pH.
Варьирование RAR обеспечило возможность выращивания растений с различными RGR s при относительно постоянных условиях электропроводности раствора и pH.
Введение
Традиционные методы культивирования в растворе часто мало напоминают условия, наблюдаемые в естественных экосистемах (Ingestad, 1982). Начнем с того, что растения истощают питательные вещества из раствора до тех пор, пока запасы не истощатся и не наступит дефицит. Таким образом, условия эксперимента со временем меняются. Кроме того, в исследованиях часто использовались концентрации питательных веществ, во много раз превышающие уровни, обнаруженные в плодородных почвах. Использование таких высоких концентраций питательных веществ создает искусственную среду для укоренения, которая потенциально может изменить модели поглощения и поглощения питательных веществ (Ingestad and Lund, 1979; Asher and Blamey, 1987).Хотя попытки имитировать доступность питательных веществ в естественной среде желательны, использование низких концентраций питательных веществ в культуре раствора создает технические трудности. Самой сложной из этих трудностей была разработка методов, обеспечивающих растениям доступ к экспоненциально растущим количествам необходимых питательных веществ при сохранении низких концентраций питательных веществ и проводимости раствора, которые обычно наблюдаются в почвенных растворах. Дополнительным вызовом является поддержание стабильных условий (обеспечение питательными веществами, рН, электропроводность) в плохо буферных системах роста, в которых отсутствует твердая фаза.
Самой сложной из этих трудностей была разработка методов, обеспечивающих растениям доступ к экспоненциально растущим количествам необходимых питательных веществ при сохранении низких концентраций питательных веществ и проводимости раствора, которые обычно наблюдаются в почвенных растворах. Дополнительным вызовом является поддержание стабильных условий (обеспечение питательными веществами, рН, электропроводность) в плохо буферных системах роста, в которых отсутствует твердая фаза.
Для решения этих проблем был разработан ряд методов. Метод культивирования в проточном растворе (Asher et al. , 1965) и метод относительной скорости добавления (RAR) (Ingestad, 1981, 1982) получили наибольшее внимание на сегодняшний день. Техника культивирования в проточном растворе обеспечивает растениям концентрации питательных веществ, которые напоминают концентрации почвенного раствора, и облегчает поддержание постоянной концентрации тестовых ионов (с частым анализом и добавками; Asher et al. , 1965). При использовании этого метода возникло несколько проблем, в том числе его трудоемкий характер и потребность в больших объемах растворов. Кроме того, высокая скорость потока раствора требуется для удовлетворения требований экспоненциального роста (Ingestad, 1982) и предотвращения истощения всех основных питательных веществ, включая исследуемые питательные вещества (Asher et al. , 1965; Asher, 1981).
, 1965). При использовании этого метода возникло несколько проблем, в том числе его трудоемкий характер и потребность в больших объемах растворов. Кроме того, высокая скорость потока раствора требуется для удовлетворения требований экспоненциального роста (Ingestad, 1982) и предотвращения истощения всех основных питательных веществ, включая исследуемые питательные вещества (Asher et al. , 1965; Asher, 1981).
Экспоненциальный рост можно также поддерживать путем частого добавления питательных веществ в экспериментальные растворы (метод RAR (Ingestad, 1981, 1982) или запрограммированного добавления питательных веществ (Asher and Blamey, 1987)).Поскольку растворы не меняются в течение экспериментального периода, этот метод требует меньших объемов воды и меньшего количества питательных веществ, чем проточные системы культивирования растворов. Если экспоненциальное снабжение питательными веществами (RAR) меньше, чем требуется для поддержания максимальной относительной скорости роста ( RGR max ), растение RGR снизится примерно до уровня RAR (Ingestad and Lund, 1986). Таким образом, этот метод обеспечивает средства для выращивания растений с достаточным или дефицитным содержанием питательных веществ со стабильным внутренним питательным статусом.
Таким образом, этот метод обеспечивает средства для выращивания растений с достаточным или дефицитным содержанием питательных веществ со стабильным внутренним питательным статусом.
Исторически сложилось так, что метод RAR включал использование аэропонных систем доставки. Метод RAR был недавно адаптирован для использования в системах культивирования в растворе, где корни растений постоянно погружаются в экспериментальные растворы (Stadt et al. , 1992). Модифицированный метод обладает многими преимуществами, доступными при использовании аэропонных систем, включая способность поддерживать рост растений в условиях низкой ионной силы и электропроводности (EC <150 мкСм см -1 ).{{-}}\)
) в ростовых растворах потребление катионов и анионов можно сбалансировать без изменения общего соотношения питательных веществ (Ingestad and Lund, 1986; Stadt et al. , 1992). Ранее было показано, что модифицированный метод RAR можно успешно использовать при ручном внесении питательных веществ (Stadt et al. , 1992; Taylor et al. , 1998). В этой работе была разработана управляемая компьютером система доставки питательных веществ, чтобы уменьшить рабочую нагрузку и облегчить более частое добавление питательных веществ (4 раза в день), чем это возможно при добавлении вручную.{\mathrm{RGR}(\mathrm{t}{-}\mathrm{t}_{0})}\]
, 1992; Taylor et al. , 1998). В этой работе была разработана управляемая компьютером система доставки питательных веществ, чтобы уменьшить рабочую нагрузку и облегчить более частое добавление питательных веществ (4 раза в день), чем это возможно при добавлении вручную.{\mathrm{RGR}(\mathrm{t}{-}\mathrm{t}_{0})}\]
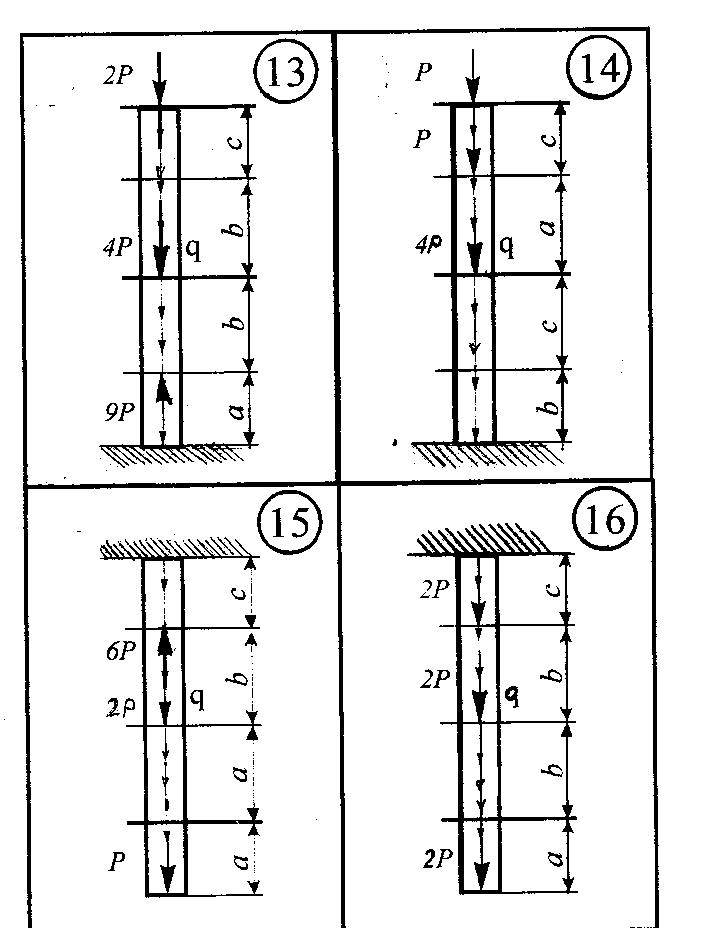 , 1992). Обратите внимание, что единицы для RAR правильно описаны как г питательного вещества (г питательного вещества для растений) -1 d -1 , а единицы для RGR правильно описаны как г г -1 растение d -1 .Для простоты при последующем цитировании RAR и RGR s будут использоваться численно идентичные единицы d -1 .
, 1992). Обратите внимание, что единицы для RAR правильно описаны как г питательного вещества (г питательного вещества для растений) -1 d -1 , а единицы для RGR правильно описаны как г г -1 растение d -1 .Для простоты при последующем цитировании RAR и RGR s будут использоваться численно идентичные единицы d -1 .Система доставки питательных веществ с компьютерным управлением
Система доставки питательных веществ с компьютерным управлением была разработана и построена для обеспечения частой (1–4 раза в день −1 ) и точной подачи питательных веществ в экспериментальные контейнеры ‡ . Аппаратная часть системы состоит из гидравлического тракта и электронно-аналоговой системы сигнализации (рис.1). Гидравлический тракт включает два многоканальных перистальтических насоса Watson Marlow (модель 202S), каждый из которых оснащен 15 приводными кассетами. Каждая из 30 приводных кассет регулирует подачу одного раствора в серию из шести из 180 электрических трехходовых пережимных клапанов (сгруппированных в три комплекта по 60). Десять таких серий (всего 60 клапанов) регулируют подачу одного питательного раствора в 60 экспериментальных емкостей. На рис. 1 представлена схема, иллюстрирующая поток питательного раствора из одного из трех резервуаров с питательным раствором через первую приводную кассету на каждом насосе к двум (из 30) наборам из шести клапанов, а затем в экспериментальные контейнеры.Когда он открыт (показан жирными стрелками на рис. 1), один клапан регулирует поток одного раствора в один контейнер для выращивания. Когда все шесть клапанов в серии клапанов закрыты (сплошная линия), питательный раствор циркулирует обратно в резервуар с питательным веществом. Система позволяет независимо поставлять три отдельных решения в каждый контейнер для выращивания.
Каждая из 30 приводных кассет регулирует подачу одного раствора в серию из шести из 180 электрических трехходовых пережимных клапанов (сгруппированных в три комплекта по 60). Десять таких серий (всего 60 клапанов) регулируют подачу одного питательного раствора в 60 экспериментальных емкостей. На рис. 1 представлена схема, иллюстрирующая поток питательного раствора из одного из трех резервуаров с питательным раствором через первую приводную кассету на каждом насосе к двум (из 30) наборам из шести клапанов, а затем в экспериментальные контейнеры.Когда он открыт (показан жирными стрелками на рис. 1), один клапан регулирует поток одного раствора в один контейнер для выращивания. Когда все шесть клапанов в серии клапанов закрыты (сплошная линия), питательный раствор циркулирует обратно в резервуар с питательным веществом. Система позволяет независимо поставлять три отдельных решения в каждый контейнер для выращивания.
Рис. 1.
Схема управляемой компьютером системы доставки питательных веществ, иллюстрирующая поток растворов для доставки питательных веществ из трех резервуаров с питательными веществами (обозначенных как растворы 1–3) через 30 приводных кассет (по 10 кассет на раствор для доставки) на два перистальтических насоса, 180 трехходовых пережимных клапанов (жирные стрелки, клапаны открыты) и экспериментальные контейнеры (незаштрихованные кружки, заштрихованные символы указывают на контейнеры, получающие питательные вещества из открытых клапанов). Каждая кассета привода насоса обеспечивает единое решение для серии из шести клапанов. Для наглядности показаны только два из 30 комплектов приводных кассет и клапанов. Если все шесть клапанов в серии закрыты, растворы для доставки возвращаются в резервуары с питательными веществами (жирная линия).
Каждая кассета привода насоса обеспечивает единое решение для серии из шести клапанов. Для наглядности показаны только два из 30 комплектов приводных кассет и клапанов. Если все шесть клапанов в серии закрыты, растворы для доставки возвращаются в резервуары с питательными веществами (жирная линия).
Рис. 1.
Схема управляемой компьютером системы доставки питательных веществ, иллюстрирующая поток растворов для доставки питательных веществ из трех резервуаров с питательными веществами (обозначенных как растворы 1–3) через 30 приводных кассет (по 10 кассет на раствор для доставки) на двух перистальтических насосах, на 180 трехходовых пережимных клапанов (жирные стрелки, клапаны открыты) и на экспериментальных контейнерах (открытые кружки, обратите внимание, заштрихованные символы указывают на контейнеры, получающие питательные вещества из открытых клапанов).Каждая кассета привода насоса обеспечивает единое решение для серии из шести клапанов. Для наглядности показаны только два из 30 комплектов приводных кассет и клапанов. Если все шесть клапанов в серии закрыты, растворы для доставки возвращаются в резервуары с питательными веществами (жирная линия).
Если все шесть клапанов в серии закрыты, растворы для доставки возвращаются в резервуары с питательными веществами (жирная линия).
Программное обеспечение RAR управляет открытием и закрытием электрических клапанов с помощью электронной/аналоговой системы сигнализации. Программа рассчитывает добавки питательных веществ ( A t , как в уравнении 3) на основе предоставленной пользователем информации, характерной для каждого эксперимента (включая W 0 , содержание питательных веществ в растениях, RGR , концентрацию питательных веществ в растворах для доставки). , и расход каждой кассеты насоса) и вычисляет время открытия и закрытия для каждого из 180 значений.Когда требуется открыть клапаны, компьютер посылает цифровые сигналы на серию из 180 релейных переключателей, которые преобразуют цифровые сигналы в аналоговые. Реле переключается, затем активирует определенные клапаны. Когда на клапаны подается питание, они открываются, позволяя питательным растворам циркулировать в емкости для роста. Питательные добавки вносят один раз в день, если время открытия клапана не превышает 20 минут, и в это время добавки делят на четыре аликвоты в течение 24 часов.
Питательные добавки вносят один раз в день, если время открытия клапана не превышает 20 минут, и в это время добавки делят на четыре аликвоты в течение 24 часов.
Экспериментальный проект
Во всех описанных здесь экспериментах использовались два основных экспериментальных плана: (i) анализ динамики во времени и (ii) факторные обработки.Во всех экспериментах, кроме последнего, питательные вещества добавляли при RAR 0,20 d -1 . Предыдущие эксперименты показали, что этот RAR был немного ниже, чем RGR max в этих экспериментальных условиях. Таким образом, рост растений должен быть ограничен RAR. Во всех экспериментах использовалась рандомизированная блочная конструкция с тремя или четырьмя статистически независимыми повторами, всего 60 контейнеров. Каждый эксперимент проводили не менее двух раз. Временные эксперименты проводились в течение 21–30 дней.Для точного определения RGR s растения собирали каждые вторые сутки в течение экспериментального периода и сушили до постоянной массы.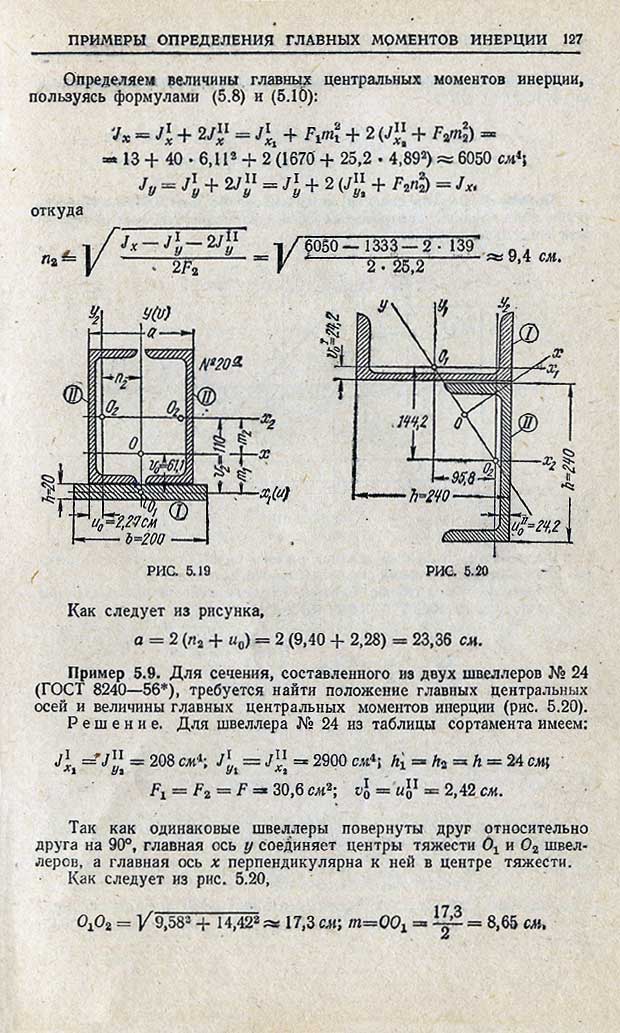 Сухие массы (г горшка -1 ) были преобразованы в натуральные логарифмы и нанесены на график зависимости от времени. Наклон наиболее подходящей линии регрессии представляет собой среднее значение RGR для нанесенных точек.
Сухие массы (г горшка -1 ) были преобразованы в натуральные логарифмы и нанесены на график зависимости от времени. Наклон наиболее подходящей линии регрессии представляет собой среднее значение RGR для нанесенных точек.
Общие методы выращивания
Питательные растворы : Пропорции питательных веществ в растворах для доставки и роста были основаны на растворах, использованных Ingestad and Stoy (1982), Pettersson and Strid (1989) и Stadt et al. (1992). В последующем тексте уровни питательных веществ в растворе выражены в виде концентрации азота. Эти растворы содержат указанную концентрацию азота, а также дополнительные питательные вещества в молярных пропорциях, указанных в таблице 1. Состав трех растворов для доставки питательных веществ (поставляемых управляемая компьютером система доставки питательных веществ) варьировались в зависимости от плана эксперимента и использовались для приготовления всех растворов для предварительной обработки и экспериментальных растворов.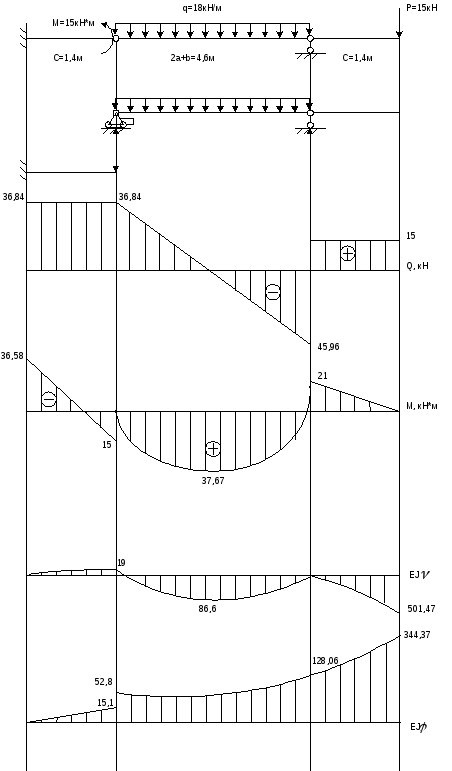 Раствор для доставки 1 содержал NH 4 NO 3 , Mg(NO 3 ) 2 .6H 2 O, Ca(NO 3 ) 2 .4H 2 O и 0,0025 г л -1 Vitavax (для ограничения роста грибов). ДОСТАВКА ДЛЯ РЕШЕНИЯ 2 CH 2 PO 4 , KNO 4 , K 2 , K 2 , MNSO 4 , MNSO 4 .H 2 O, H 3 BO 3 , ZNSO 4 .7H 2 O, CuSO 4 .5H 2 O и Na 2 MoO 4 .2H 2 O. Раствор доставки 3 содержал FeCl 0{{-}}\)
Раствор для доставки 1 содержал NH 4 NO 3 , Mg(NO 3 ) 2 .6H 2 O, Ca(NO 3 ) 2 .4H 2 O и 0,0025 г л -1 Vitavax (для ограничения роста грибов). ДОСТАВКА ДЛЯ РЕШЕНИЯ 2 CH 2 PO 4 , KNO 4 , K 2 , K 2 , MNSO 4 , MNSO 4 .H 2 O, H 3 BO 3 , ZNSO 4 .7H 2 O, CuSO 4 .5H 2 O и Na 2 MoO 4 .2H 2 O. Раствор доставки 3 содержал FeCl 0{{-}}\)
Доли питательных веществ (мол. % по отношению к N) в фоновых растворах и растворах для доставки питательных веществ
Элемент . 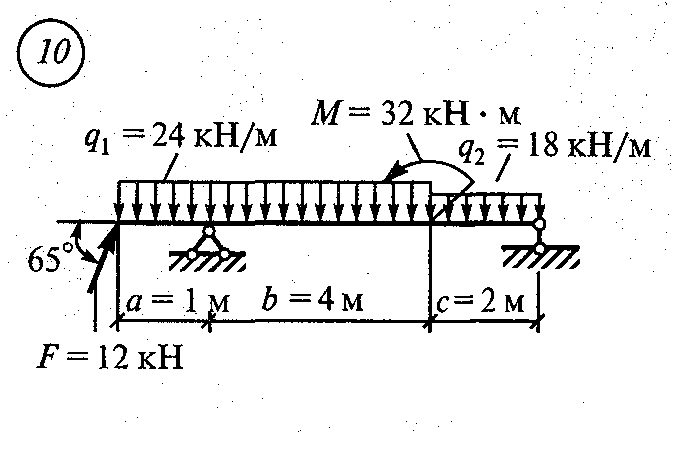 | Концентрация (мол. %) относительно N .{{-}} \) | 75 | 64 | |
|---|---|---|---|---|
| фосфористой | 6,7 | 6,7 | ||
| калия | 30,7 | 30,7 | ||
| кальция | 7.1 | 7.1 | ||
| Magnesium | 5.7 | 5.7 | 5.7 | |
| 3.0 | 3.0 | 3.0 | ||
| Iron | 1,8 × 10 -1 | 1.8 × 10 -1 8 | ||
| 1,0 × 10 -1 | 1,0 × 10 -1 | |||
| Boron | 2,6 × 10 -1 | 2,6 × 10 -1 | ||
| Zinc | 1.3 × 10 -2 | 1,3 × 10 -2 | ||
| медь | 5.7 × 10 -3 | 5.7 × 10 -3 | ||
| Молибден | 1. {{-}} \) {{-}} \) | 75 | 64 | |
| фосфористой | 6,7 | 6,7 | ||
| калия | 30,7 | 30,7 | ||
| кальция | 7.1 | 7.1 | ||
| Magnesium | 5.7 | 5.7 | 5.7 | |
| 3.0 | 3.0 | 3.0 | ||
| Iron | 1,8 × 10 -1 | 1.8 × 10 -1 8 | ||
| 1,0 × 10 -1 | 1,0 × 10 -1 | |||
| Boron | 2,6 × 10 -1 | 2,6 × 10 -1 | ||
| Zinc | 1.3 × 10 -2 | 1,3 × 10 -2 | ||
| медь | 5.7 × 10 -3 | 5.7 × 10 -3 | ||
| Молибден | 1.0 × 10 -3 9043 -3 | 1,0 × 10 -3 | ||
5. 4 × 10 -1 4 × 10 -1 | ||||
Пропорции питательных веществ ( мол.% по отношению к N) в фоновых растворах и растворах для доставки питательных веществ
Элемент . | Концентрация (мол. %) относительно N . | .{{-}} \) | 75 | 64 | |
|---|---|---|---|---|---|
| фосфористой | 6,7 | 6,7 | |||
| калия | 30,7 | 30,7 | |||
| кальция | 7.1 | 7.1 | |||
| Magnesium | 5.7 | 5.7 | 5.7 | ||
| 3.0 | 3.0 | 3.0 | |||
| Iron | 1,8 × 10 -1 | 1.8 × 10 -1 8 | |||
| 1,0 × 10 -1 | 1,0 × 10 -1 | ||||
| Boron | 2,6 × 10 -1 | 2,6 × 10 -1 | |||
| Zinc | 1. 3 × 10 -2 3 × 10 -2 | 1,3 × 10 -2 | |||
| медь | 5.7 × 10 -3 | 5.7 × 10 -3 | |||
| Молибден | 1.{{-}} \) | 75 | 64 | ||
| фосфористой | 6,7 | 6,7 | |||
| калия | 30,7 | 30,7 | |||
| кальция | 7.1 | 7.1 | |||
| Magnesium | 5.7 | 5.7 | 5.7 | ||
| 3.0 | 3.0 | 3.0 | |||
| Iron | 1,8 × 10 -1 | 1.8 × 10 -1 8 | |||
| 1,0 × 10 -1 | 1,0 × 10 -1 | ||||
| Boron | 2,6 × 10 -1 | 2,6 × 10 -1 | |||
| Zinc | 1.3 × 10 -2 | 1,3 × 10 -2 | |||
| медь | 5. 7 × 10 -3 7 × 10 -3 | 5.7 × 10 -3 | |||
| Молибден | 1.0 × 10 -3 | 1,0 × 10 -3 | |||
| 5.4 × 10 -1 | |||||
Подготовка растительного материала (период предварительной обработки ) : Семена Triticum aestivum L. cv. Katepwa стерилизовали поверхность в 1,1% растворе гипохлорита натрия (об./об.) в течение 20 мин и проращивали в течение ночи в аэрированном растворе, содержащем 0,005 г л -1 Vitavax (Uniroyal Chemical Ltd., Калгари, AB, Канада) для ограничения роста грибков. Затем семена выращивали в аквариумах (контейнеры из плексигласа 20 л; по 300 семян в каждом) на нейлоновой сетке, подвешенной над аэрированным раствором, содержащим фоновые питательные вещества (50, 200 или 500 мкМ N, в зависимости от схемы эксперимента), доведенным до pH 4,3 с помощью 1,0 или 0,1. М HCl. Всходы прореживали через 3 дня до 150 растений на аквариум. В это время 12 проростков высушивали в течение 2 ч (до постоянного веса) при 60 °C для определения исходного сухого веса ( W 0 ) для расчета добавок питательных веществ на оставшуюся часть периода предварительной обработки.Добавление питательных веществ за каждый день ( A t ) рассчитывали по уравнению 3. Расчетные добавки питательных веществ для периода предварительной обработки были уменьшены наполовину в экспериментах, представленных ниже на рис. 6–10, поскольку запасы семян обеспечивали проростки достаточным количеством питательных веществ. для ускорения RGR выше желаемого уровня. Начиная примерно через 3–4 дня после прорастания (как только проростки становятся достаточно большими, чтобы их можно было отделить от семян), каждый день собирали подвыборки ( n = 8) растений, сушили и взвешивали (без семян) для определения роста в долечебный период.Сухая масса растений в последний день периода предварительной обработки использовалась для оценки W 0 для расчета добавок питательных веществ на экспериментальный период.
М HCl. Всходы прореживали через 3 дня до 150 растений на аквариум. В это время 12 проростков высушивали в течение 2 ч (до постоянного веса) при 60 °C для определения исходного сухого веса ( W 0 ) для расчета добавок питательных веществ на оставшуюся часть периода предварительной обработки.Добавление питательных веществ за каждый день ( A t ) рассчитывали по уравнению 3. Расчетные добавки питательных веществ для периода предварительной обработки были уменьшены наполовину в экспериментах, представленных ниже на рис. 6–10, поскольку запасы семян обеспечивали проростки достаточным количеством питательных веществ. для ускорения RGR выше желаемого уровня. Начиная примерно через 3–4 дня после прорастания (как только проростки становятся достаточно большими, чтобы их можно было отделить от семян), каждый день собирали подвыборки ( n = 8) растений, сушили и взвешивали (без семян) для определения роста в долечебный период.Сухая масса растений в последний день периода предварительной обработки использовалась для оценки W 0 для расчета добавок питательных веществ на экспериментальный период.
Экспериментальный период : Через 9 дней отработанные семена удаляли со всех растений и восемь однородных проростков переносили в каждый из 60 полиэтиленовых контейнеров, наполненных 10 л аэрируемого фонового раствора. Растворы доводили до рН 4,3. Эти фоновые растворы готовили с помощью компьютерного управления подачей соответствующего объема растворов для доставки питательных веществ в 10 л дистиллированной воды для достижения желаемой фоновой концентрации (50–1000 мкМ N).После этого питательные вещества поставлялись в экспоненциально возрастающих количествах, в 1-4 раза d -1 . Саженцы устанавливали на непрозрачные крышки из плексигласа, которые помещали поверх контейнеров, чтобы ограничить рост водорослей. К питательным растворам периодически добавляли дистиллированную воду, чтобы компенсировать потери воды на испарение и транспирацию. Контейнеры подвешивали в общей водяной бане, чтобы ограничить колебания температуры и поддерживать постоянную температуру во всех экспериментальных растворах.
pH раствора и электропроводность периодически контролировались с помощью pH-метра Radiometer PHM80 и измерителя электропроводности Radiometer CDM80 с зондом CDC104. Измерения проводились перед посадкой, три раза в неделю (непосредственно перед внесением питательных веществ) и сразу после сбора урожая.
Эксперименты проводились в двух ростовых камерах с контролируемой средой, при 16-часовом освещении и 8-часовом темноте. Температуры светового периода колебались в пределах 20–24 °С, а темного – 16,7–19,5 °С.Относительная влажность колебалась от 50% до 84% в светлое время суток и от 75% до 100% в темное время суток. Температура раствора варьировала от 19°С до 23°С в световой период и от 18°С до 21°С в темное время суток. Вегетационная камера освещалась 103 холодными белыми люминесцентными лампами (25 Вт) и 16 лампами накаливания (150 Вт), расположенными на высоте 1,3 м над основанием растений. Средняя плотность фотосинтетического потока фотонов ( PPFD ) для одиночных экспериментов, проведенных в обеих камерах, колебалась от 332 до 471 мкмоль·м −2 с −1 . {{+}}\)
{{+}}\)
Фоновые концентрации питательных веществ : Этот эксперимент был разработан, чтобы определить, повлияет ли начальная фоновая концентрация питательных веществ на скорость роста растений. В эксперименте использовали фон 200 мкМ N для периода предварительной обработки и 15 фоновых концентраций (50, 100, 150, 200, 250, 300, 350, 400, 450, 500, 600, 700, 800, 900 и 1000 мкМ N). за 21-дневный экспериментальный период.Все питательные вещества подавались в тех же массовых пропорциях, что и в предыдущем эксперименте (таблица 1), при RAR 0,20 d -1 .
Динамика роста во времени (фон 500 мкМ N) : Этот эксперимент был использован для определения возможности поддержания 0,20 сут -1 RGR при фоновой концентрации 500 мкМ азота и 0,20 сут -1 RAR в течение 22-дневного экспериментального периода. Никаких изменений в пропорции питательных веществ не вносилось.
Никаких изменений в пропорции питательных веществ не вносилось.
Фон кальция : Этот эксперимент был разработан, чтобы определить, позволит ли увеличение концентрации кальция в фоновых растворах снизить фоновые уровни других питательных веществ (т.е. с использованием фона N 200 вместо 500 мкМ). Кальций добавляли из исходного раствора 1,0 М CaCl 2 к фоновому 200 мкМ N, чтобы довести конечную концентрацию Ca до 14, 95, 195, 400, 1000 и 2000 мкМ. Растения выращивали в течение 9 дней при RAR 0,20 дня -1 .
Фоновые концентрации питательных веществ (400 мкМ общего кальция) : Этот 21-дневный эксперимент был разработан для определения того, будет ли общая концентрация 400 мкМ Са в растворах предварительной обработки (200 мкМ N) и экспериментальных фоновых растворах (50–1000 мкМ N) позволяют снизить уровень других фоновых питательных веществ.{{+}}\)
). Временная динамика для различных RAR : Этот 24-дневный эксперимент был использован для определения того, обеспечит ли изменение RAR питательных веществ контроль RGR и повлияет ли оно на вызванные растениями изменения в растворе EC и pH.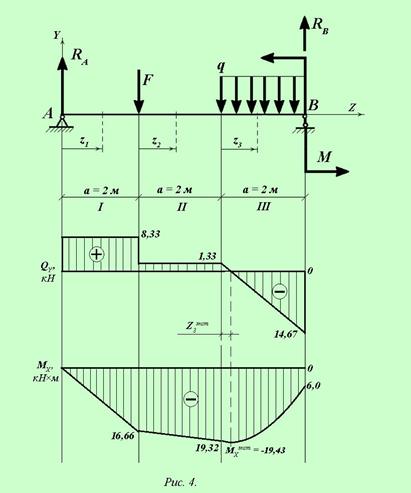 Фон 50 мкМ N плюс 400 мкМ CaCl 2 использовали для предварительной обработки и экспериментального периода. Питательные вещества подавались при RAR 0,09, 0,12, 0,15, 0,18 и 0,21 дня -1 как в растворах для предварительной обработки, так и в экспериментальных растворах.
Фон 50 мкМ N плюс 400 мкМ CaCl 2 использовали для предварительной обработки и экспериментального периода. Питательные вещества подавались при RAR 0,09, 0,12, 0,15, 0,18 и 0,21 дня -1 как в растворах для предварительной обработки, так и в экспериментальных растворах.
Результаты и обсуждение
Динамика роста во времени (фон 200 мкМ N)
RGR растений в период предварительной обработки (0.305 д -1 ; рис. 2А, незаштрихованные символы) превысило 0,20 d -1 RAR. Эта более высокая скорость роста, по-видимому, была связана с мобилизацией запасов питательных веществ из семян, которые снабжали растения питательными веществами, превышающими те, которые поставлялись с питательными добавками. Рост был логарифмически линейным в течение первых 14 дней экспериментального периода, но RGR (0,084 дня -1 ) был ниже, чем RAR (0,20 дня -1 ).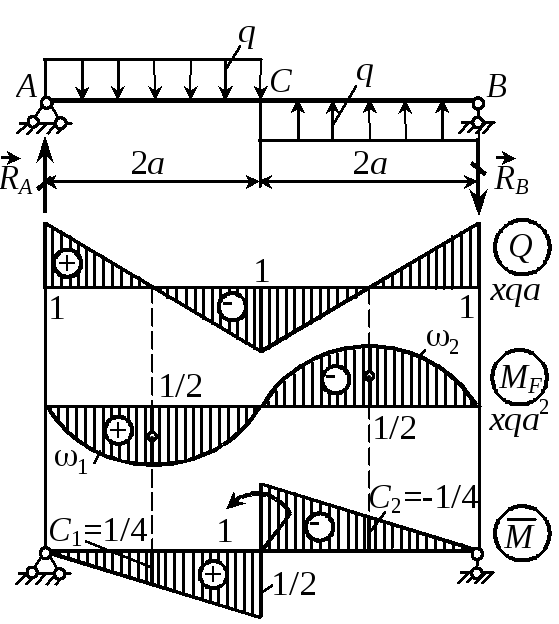 Обратите внимание на различия между ожидаемым ростом (штриховая линия) и наблюдаемым ростом (сплошная линия) на рис.2А. RGR впоследствии ускорился до 0,171 д -1 до конца экспериментального периода (рис. 2А). Ограничение роста в течение первых 14 дней эксперимента сопровождалось видимыми признаками стресса. Корни приобретают светло-коричневую окраску с ограниченным развитием боковых корней. Рост боковых корней возобновился после 14-го дня, и новый рост выглядел белым и здоровым до конца экспериментального периода. Снижение видимых признаков стресса совпало с более чем двукратным увеличением RGR (рис.{{+}}\)
Обратите внимание на различия между ожидаемым ростом (штриховая линия) и наблюдаемым ростом (сплошная линия) на рис.2А. RGR впоследствии ускорился до 0,171 д -1 до конца экспериментального периода (рис. 2А). Ограничение роста в течение первых 14 дней эксперимента сопровождалось видимыми признаками стресса. Корни приобретают светло-коричневую окраску с ограниченным развитием боковых корней. Рост боковых корней возобновился после 14-го дня, и новый рост выглядел белым и здоровым до конца экспериментального периода. Снижение видимых признаков стресса совпало с более чем двукратным увеличением RGR (рис.{{+}}\)
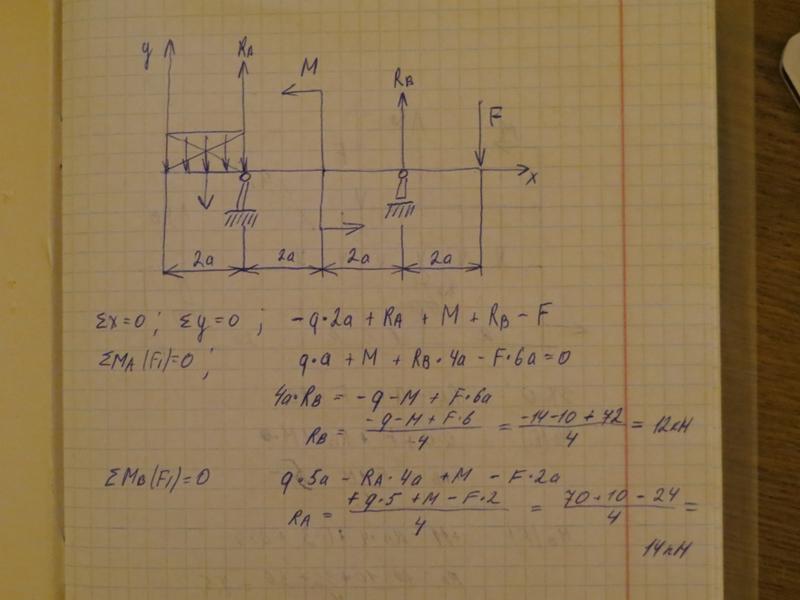 Закрытые символы обозначают рост в течение экспериментального периода. Различное затенение символов в течение экспериментального периода указывает на изменение скорости роста растений.{{+}}\) ) фонового раствора за 30-дневный экспериментальный период. Изменения в растворе EC (B) и pH (C) измеряли три раза в неделю. Сухой вес растений (г горшка -1 ) был логарифмически преобразован и нанесен на график с линией регрессии первого порядка. Значения являются средними ±SE ( n =4). Открытые символы обозначают рост растений в период предварительной обработки. Закрытые символы обозначают рост в течение экспериментального периода. Различное затенение символов в течение экспериментального периода указывает на изменение скорости роста растений.Пунктирная линия, наиболее подходящая регрессия для периода до лечения. Сплошная линия, наиболее точное соответствие регрессии для экспериментального периода. Пунктирная линия представляет прогнозируемый рост для RGR на 0,20 дня -1 .
Закрытые символы обозначают рост в течение экспериментального периода. Различное затенение символов в течение экспериментального периода указывает на изменение скорости роста растений.{{+}}\) ) фонового раствора за 30-дневный экспериментальный период. Изменения в растворе EC (B) и pH (C) измеряли три раза в неделю. Сухой вес растений (г горшка -1 ) был логарифмически преобразован и нанесен на график с линией регрессии первого порядка. Значения являются средними ±SE ( n =4). Открытые символы обозначают рост растений в период предварительной обработки. Закрытые символы обозначают рост в течение экспериментального периода. Различное затенение символов в течение экспериментального периода указывает на изменение скорости роста растений.Пунктирная линия, наиболее подходящая регрессия для периода до лечения. Сплошная линия, наиболее точное соответствие регрессии для экспериментального периода. Пунктирная линия представляет прогнозируемый рост для RGR на 0,20 дня -1 .
Поскольку в методе RAR используются предварительно запрограммированные добавки питательных веществ, снижение роста ( RGR
Видимые признаки стресса в корнях, подобные описанным здесь, ранее интерпретировались как отражение периода приспособления растений к изменениям внутреннего статуса питательных веществ, которые следуют за началом добавления питательных веществ с помощью метода RAR (Ingestad, 1981).Низкие темпы роста ( RGR  Интересно отметить, что Stadt et al. (1992) наблюдали симптомы питательного стресса у растений, выращенных при низких фоновых уровнях питательных веществ (0–180 мкМ N), но эти симптомы не сопровождались снижением RGR ниже RAR. Это могло отражать низкие значения RAR, использованные в их исследовании (0.05 и 0,15 д -1 ). При высоком RAR, используемом в этом исследовании (0,20 d -1 ), недостаточное снабжение питательными веществами в фоновых растворах могло создать реальные ограничения для роста в течение экспериментального периода. Обратите внимание, что неадекватная фоновая концентрация в период предварительной обработки не обязательно приведет к снижению роста, поскольку мобилизация запасов семян может обеспечить достаточное количество питательных веществ, необходимых для здорового роста. Это подтверждается скоростью роста 0,305 дня -1 , наблюдаемой в период до обработки.
Интересно отметить, что Stadt et al. (1992) наблюдали симптомы питательного стресса у растений, выращенных при низких фоновых уровнях питательных веществ (0–180 мкМ N), но эти симптомы не сопровождались снижением RGR ниже RAR. Это могло отражать низкие значения RAR, использованные в их исследовании (0.05 и 0,15 д -1 ). При высоком RAR, используемом в этом исследовании (0,20 d -1 ), недостаточное снабжение питательными веществами в фоновых растворах могло создать реальные ограничения для роста в течение экспериментального периода. Обратите внимание, что неадекватная фоновая концентрация в период предварительной обработки не обязательно приведет к снижению роста, поскольку мобилизация запасов семян может обеспечить достаточное количество питательных веществ, необходимых для здорового роста. Это подтверждается скоростью роста 0,305 дня -1 , наблюдаемой в период до обработки.
Фоновые концентрации питательных веществ
В этом эксперименте манипулировали фоновыми концентрациями питательных веществ, чтобы преодолеть возможное ограничение по питательным веществам. При повышении фоновых концентраций с 50 мкМ N до 500 мкМ N увеличивалось накопление растительной биомассы (рис. 3), а общий RGR увеличивался с 0,08 до 0,165 d -1 . Небольшой дополнительный рост наблюдался при концентрациях более 500 мкМ N (рис.3). Ingestad (1972) обнаружил, что рост Cucumis sativus L. (огурец) снижался, когда концентрации питательных веществ снижались ниже 14 мМ N. В другом исследовании Ingestad and Lund (1979) пришли к выводу, что общая концентрация N должна быть не менее 1,8 мМ. но не более 19,3 мМ для максимального роста Betula verrucosa Ehrh. (береза). Эти более высокие оптимальные концентрации могли отражать различия в экспериментальных видах (пшеница по сравнению с огурцом или березой) или методах (культивирование в растворе по сравнению с культивированием в питательном тумане).Штадт и др. (1992) сообщают о линейной реакции роста пшеницы при фоновых концентрациях в диапазоне от 0 до 360 мкМ N и RAR 0,15 дня -1 .
При повышении фоновых концентраций с 50 мкМ N до 500 мкМ N увеличивалось накопление растительной биомассы (рис. 3), а общий RGR увеличивался с 0,08 до 0,165 d -1 . Небольшой дополнительный рост наблюдался при концентрациях более 500 мкМ N (рис.3). Ingestad (1972) обнаружил, что рост Cucumis sativus L. (огурец) снижался, когда концентрации питательных веществ снижались ниже 14 мМ N. В другом исследовании Ingestad and Lund (1979) пришли к выводу, что общая концентрация N должна быть не менее 1,8 мМ. но не более 19,3 мМ для максимального роста Betula verrucosa Ehrh. (береза). Эти более высокие оптимальные концентрации могли отражать различия в экспериментальных видах (пшеница по сравнению с огурцом или березой) или методах (культивирование в растворе по сравнению с культивированием в питательном тумане).Штадт и др. (1992) сообщают о линейной реакции роста пшеницы при фоновых концентрациях в диапазоне от 0 до 360 мкМ N и RAR 0,15 дня -1 . {{+}}\)
{{+}}\)
Динамика роста во времени (фон 500 мкМ N)
Когда фоновая концентрация питательных веществ была увеличена до 500 мкМ N, растение RGR оставалось близким к 0,20 дня -1 RAR в течение 22 дней экспериментального периода (рис. 4А). Видимые признаки стресса (снижение бокового роста, коричневая окраска) все еще наблюдались в течение первой недели роста, но эти признаки исчезали после 8-го дня, когда возобновлялся рост боковых корней.В отличие от предыдущего эксперимента с временной динамикой, где RGR < RAR и EC раствора увеличилась до ~1300 мкСм см -1 (рис. 2), EC раствора оставалась практически постоянной (40–90 мкСм см -1 ). , рис. 4Б). Таким образом, оказалось, что поглощение питательных веществ находится в равновесии с поступлением питательных веществ. Тем не менее, pH ростового раствора через 2 недели увеличился до более чем pH 6,5 (рис. 4C), что свидетельствует о том, что поглощение анионов превышает поглощение катионов.
Тем не менее, pH ростового раствора через 2 недели увеличился до более чем pH 6,5 (рис. 4C), что свидетельствует о том, что поглощение анионов превышает поглощение катионов.
Рис.{{+}}\)
) фонового раствора за 22-дневный экспериментальный период. Изменения в растворе EC (B) и pH (C) измеряли три раза в неделю. Сухой вес растений (г горшка -1 ) был логарифмически преобразован и нанесен на график с линией регрессии первого порядка. Значения являются средними ±SE ( n =4). Открытые символы обозначают рост растений в период предварительной обработки. Закрытые символы обозначают рост в течение экспериментального периода. Пунктирная линия, наиболее подходящая регрессия для периода до лечения. Сплошная линия, наиболее точное соответствие регрессии для экспериментального периода.{{+}}\) ) фонового раствора за 22-дневный экспериментальный период. Изменения в растворе EC (B) и pH (C) измеряли три раза в неделю. Сухой вес растений (г горшка -1 ) был логарифмически преобразован и нанесен на график с линией регрессии первого порядка. Значения являются средними ±SE ( n =4). Открытые символы обозначают рост растений в период предварительной обработки. Закрытые символы обозначают рост в течение экспериментального периода. Пунктирная линия, наиболее подходящая регрессия для периода до лечения. Сплошная линия, наиболее точное соответствие регрессии для экспериментального периода.Пунктирная линия представляет прогнозируемый рост для RGR на 0,20 дня -1 .
Значения являются средними ±SE ( n =4). Открытые символы обозначают рост растений в период предварительной обработки. Закрытые символы обозначают рост в течение экспериментального периода. Пунктирная линия, наиболее подходящая регрессия для периода до лечения. Сплошная линия, наиболее точное соответствие регрессии для экспериментального периода.Пунктирная линия представляет прогнозируемый рост для RGR на 0,20 дня -1 .Несмотря на то, что логарифмический линейный рост был достигнут при использовании фоновой концентрации азота 500 мкМ (рис. 4А), симптомы стресса, наблюдаемые в течение первой недели эксперимента, свидетельствовали о продолжающемся дефиците некоторых основных питательных веществ. Коричневое обесцвечивание корней и уменьшение роста боковых побегов напоминали симптомы, связанные с дефицитом кальция (Loneragan et al. , 1969).
Фон кальция
Чтобы определить, можно ли облегчить симптомы стресса, наблюдавшиеся в предыдущем эксперименте, и можно ли снизить уровни фоновых питательных веществ за счет увеличения поступления кальция, общие концентрации кальция в 200 мкМ фоновых растворах N варьировали от 14 до 2000 мкМ. {{+}}\)
{{+}}\)
Фоновые питательные вещества плюс 400 мкМ общего кальция
Этот эксперимент был разработан, чтобы определить, можно ли снизить фоновые концентрации других питательных веществ при использовании более высокого фонового уровня кальция (400 мкМ). Когда другие фоновые питательные вещества варьировались в диапазоне от 50 до 1000 мкМ N, рост увеличивался линейно (рис.{{+}}\)
) и 400 мкМ общего Ca за 21-дневный экспериментальный период. Изменения в растворе EC (B) и pH (C) измеряли три раза в неделю. Значения являются средними ±SE ( n =4). EC ростовых растворов мало изменилась в течение экспериментального периода для всех фоновых обработок. Тем не менее, значения EC при более низких фоновых обработках (50–200 мкМ N) демонстрировали меньшие изменения во времени, чем значения при более высоких фоновых обработках. Для более высоких фоновых обработок (700–1000 мкМ N) наблюдалась большая тенденция к снижению показателей ЕС между 9 и 19 днями (рис.6B), предполагая, что RGR растений при этих обработках с высоким фоном был выше, чем при обработках с низким фоном N, и больше, чем RAR. Это подтвердилось при сравнении общих темпов роста. Относительная скорость роста при обработках с высоким фоном была на 0,005-0,015 г -1 растение d -1 больше, чем при обработках 50-200 мкМ азота (скорости варьировались от 0,176 до 0,181 г -1 растение d — 1 ). Во всех фоновых обработках рН увеличился с 4.3 до более чем 7,0 (фиг. 6C), что еще раз свидетельствует о том, что поглощение анионов превышает поглощение катионов.
Для более высоких фоновых обработок (700–1000 мкМ N) наблюдалась большая тенденция к снижению показателей ЕС между 9 и 19 днями (рис.6B), предполагая, что RGR растений при этих обработках с высоким фоном был выше, чем при обработках с низким фоном N, и больше, чем RAR. Это подтвердилось при сравнении общих темпов роста. Относительная скорость роста при обработках с высоким фоном была на 0,005-0,015 г -1 растение d -1 больше, чем при обработках 50-200 мкМ азота (скорости варьировались от 0,176 до 0,181 г -1 растение d — 1 ). Во всех фоновых обработках рН увеличился с 4.3 до более чем 7,0 (фиг. 6C), что еще раз свидетельствует о том, что поглощение анионов превышает поглощение катионов.
В то время как вызванные растениями колебания рН указывали на дисбаланс в потреблении катионов/анионов, этот эксперимент подтвердил, что здоровый рост может быть достигнут при фоновом уровне 50 мкМ азота при общей концентрации 400 мкМ Са.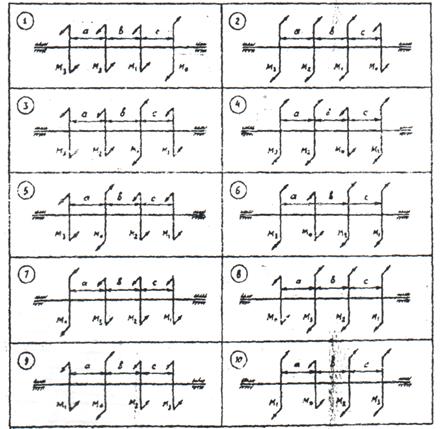 {{+}}\)
{{+}}\)
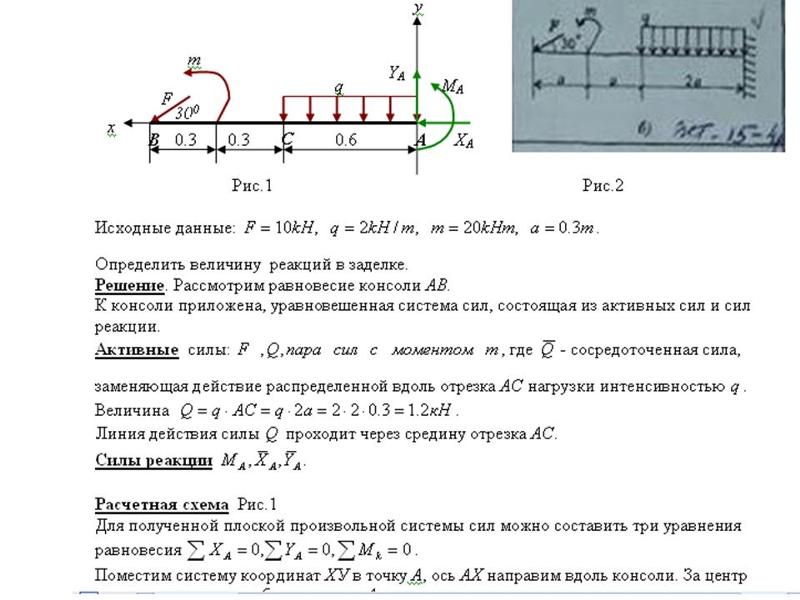 {{+}}\) ) фон и 400 мкМ всего Ca в течение 21-дневного экспериментального периода.Сплошная линия, наиболее точно соответствующая линии регрессии для роста в течение экспериментального периода. Пунктирная линия представляет собой линию регрессии для относительной скорости роста 0,20 d -1 . Изменения в растворе EC (B) и pH (C) измеряли три раза в неделю. Значения являются средними ±SE ( n =4).
{{+}}\) ) фон и 400 мкМ всего Ca в течение 21-дневного экспериментального периода.Сплошная линия, наиболее точно соответствующая линии регрессии для роста в течение экспериментального периода. Пунктирная линия представляет собой линию регрессии для относительной скорости роста 0,20 d -1 . Изменения в растворе EC (B) и pH (C) измеряли три раза в неделю. Значения являются средними ±SE ( n =4).Временной курс для различных RAR
В этом заключительном эксперименте в течение 24-дневного экспериментального периода изучалось влияние различных RAR (0,09–0,21 дня -1 ) на рост растений и вызванные растениями изменения в питательном растворе.Как и ожидалось, рост растений увеличивался с увеличением скорости добавления питательных веществ (фиг. 10А), подтверждая предыдущие сообщения о том, что модифицированный метод RAR обеспечивает контроль над растением RGR (Stadt et al. , 1992). В этом эксперименте также было показано, что различные RGR могут поддерживаться при относительно скромных изменениях, вызванных растениями в ростовых растворах. При обработке с низким RAR (0,09–0,15 d -1 ) EC раствора колебалась между 125 и 185 мкСм·см -1 , а pH раствора варьировался примерно на 0.5 единиц pH в течение 24-дневного периода роста. Это контрастирует с изменениями pH более чем на три порядка в некоторых из этих экспериментов по оптимизации (см., например, рис. 6C, 7C) или в экспериментах с обычной культурой в растворе (см., например, Taylor and Foy, 1985). EC раствора показал большее увеличение при обработке 0,18 и 0,21 дня -1 RAR. Обработка 0,21 дня -1 RAR была выбрана для обеспечения условий, при которых RAR> RGR max , таким образом, ожидалось повышение ЕС.Как и в предыдущих экспериментах, где RAR > , RGR и EC раствора увеличивались, наблюдалось параллельное снижение pH раствора. Обработка 0,18 дня -1 RAR была ниже расчетной RGR max для этих экспериментальных условий. Тем не менее, рост при лечении 0,18 дня -1 RAR не был таким быстрым, как ожидалось ( RGR
При обработке с низким RAR (0,09–0,15 d -1 ) EC раствора колебалась между 125 и 185 мкСм·см -1 , а pH раствора варьировался примерно на 0.5 единиц pH в течение 24-дневного периода роста. Это контрастирует с изменениями pH более чем на три порядка в некоторых из этих экспериментов по оптимизации (см., например, рис. 6C, 7C) или в экспериментах с обычной культурой в растворе (см., например, Taylor and Foy, 1985). EC раствора показал большее увеличение при обработке 0,18 и 0,21 дня -1 RAR. Обработка 0,21 дня -1 RAR была выбрана для обеспечения условий, при которых RAR> RGR max , таким образом, ожидалось повышение ЕС.Как и в предыдущих экспериментах, где RAR > , RGR и EC раствора увеличивались, наблюдалось параллельное снижение pH раствора. Обработка 0,18 дня -1 RAR была ниже расчетной RGR max для этих экспериментальных условий. Тем не менее, рост при лечении 0,18 дня -1 RAR не был таким быстрым, как ожидалось ( RGR  Поскольку в последние 8–10 сут эксперимента в растворе начинали накапливаться питательные вещества, наблюдали изменение ЕС и рН раствора.{{+}}\)
Поскольку в последние 8–10 сут эксперимента в растворе начинали накапливаться питательные вещества, наблюдали изменение ЕС и рН раствора.{{+}}\)
Эти эксперименты демонстрируют, что управляемая компьютером система доставки питательных веществ может использоваться для управления фоновым снабжением питательными веществами, ежедневными добавками питательных веществ и балансом катионов/анионов для поддержания высоких темпов роста растений (0,2 г −1 растения d − 1 ). В то время как оптимальное снабжение питательными веществами может зависеть от генотипа, сбалансированное снабжение и потребление питательных веществ обеспечивает средства поддержания логарифмически-линейного роста при относительно постоянных условиях EC и pH раствора.
Мы хотели бы поблагодарить Кена Штадта за его ценную техническую помощь с системой доставки с компьютерным управлением. Это исследование было поддержано средствами, предоставленными Советом по естественным наукам и инженерным исследованиям Канады (NSERC) по программе исследовательских грантов (грант Discovery — индивидуальный для GJT). Финансирование управляемой компьютером системы доставки питательных веществ было предоставлено GJT из средств Программы грантов для исследовательских инструментов и инструментов NSERC.
Это исследование было поддержано средствами, предоставленными Советом по естественным наукам и инженерным исследованиям Канады (NSERC) по программе исследовательских грантов (грант Discovery — индивидуальный для GJT). Финансирование управляемой компьютером системы доставки питательных веществ было предоставлено GJT из средств Программы грантов для исследовательских инструментов и инструментов NSERC.
Каталожные номера
Ашер CJ.
1981
. Ограничение внешних концентраций микроэлементов для роста растений: использование методов культивирования в проточном растворе.Journal of Plant Nutrition
3
,163
–180.Ашер СиДжей, Блейми Ф.П.
1987
. Экспериментальный контроль питательного статуса растений с помощью запрограммированного добавления питательных веществ.Journal of Plant Nutrition
10
,1371
–1380.
Ашер С.Дж., Озанн П.Г., Лонераган Дж.Ф.
1965
. Способ управления ионной средой корней растений.Почвоведение
100
,149
–156.Финденегг ГР.
1987
. Сравнительное исследование токсичности аммония при различных постоянных значениях рН питательного раствора.Растения и почва
103
,239
–243.Ингестад Т.
1972
. Потребность рассады огурцов в минеральных веществах.Физиология растений
52
,332
–338.Ингестад Т.
1981
. Питание и рост сеянцев березы и ольхи серой в растворах с низкой электропроводностью и при различных относительных нормах внесения питательных веществ.Physiologia Plantarum
52
,454
–466.Ингестад Т.
1982
. Относительная скорость добавления и внешняя концентрация; движущие переменные, используемые в исследованиях питания растений.
Растения, клетки и окружающая среда
5
,443
–453.Ингестад Т., Лунд А.
1979
. Азотный стресс у сеянцев березы.Physiologia Plantarum
45
,137
–148.Ингестад Т., Лунд А.
1986
. Теория и методика стационарного минерального питания и роста растений.Scandinavian Journal of Forest Research
1
,439
–453.Ингестад Т., Стой В.
1982
. Минеральное питание проростков пшеницы, ржи, ячменя и овса в питательных растворах.Шведский журнал сельскохозяйственных исследований
12
,185
–192.Лонераган Дж.Ф.
1979
. Интерфейс по отношению к корневой функции и росту. В: Harley JH, Russell RS, ред. Поверхность раздела почва-корень . Нью-Йорк: Academic Press, 351–367. Лонераган Дж. Ф., Сноуболл К., Симмонс В. Дж.
Ф., Сноуболл К., Симмонс В. Дж.
1969
. Реакция растений на концентрацию кальция в растворной культуре.Австралийский журнал сельскохозяйственных исследований
19
,845
–857.Мейнард Д.Н., Баркер А.В.
1969
. Изучение устойчивости растений к аммиачному питанию.Журнал Американского общества садоводческих наук
94
,235
–239.Петтерссон С., Стрид Х.
1989
.Токсичность алюминия у двух сортов пшеницы ( Triticum aestivum L.) с разной чувствительностью к алюминию в зависимости от уровня обеспеченности питательными веществами.Шведский журнал сельскохозяйственных исследований
19
,183
–191.Stadt K, Taylor GJ, Dale MRT.
1992
. Контроль относительной скорости роста путем применения метода относительной скорости добавления к традиционной системе культивирования в растворе.
Растения и почва
142
,113
–122.Тейлор Г.Дж.
1988
. Механизмы толерантности к алюминию у Triticum aestivum L. (пшеница). V. Азотное питание, индуцированный растениями рН и толерантность к алюминию; корреляция без причинно-следственной связи?Canadian Journal of Botany
66
,694
–699.Тейлор Г.Дж., Блейми Ф.П.К., Эдвардс Д.Г.
1998
. Антагонистические и синергетические взаимодействия между алюминием и марганцем при росте Vigna unguiculata при низкой ионной силе.Physiologia plantarum
104
,183
–194.Тейлор Г.Дж., Фой К.Д.
1985
. Механизмы толерантности к алюминию у Triticum aestivum L. (пшеница). IV. Роль аммиачно-нитратного питания.Canadian Journal of Botany
63
,2181
–2186.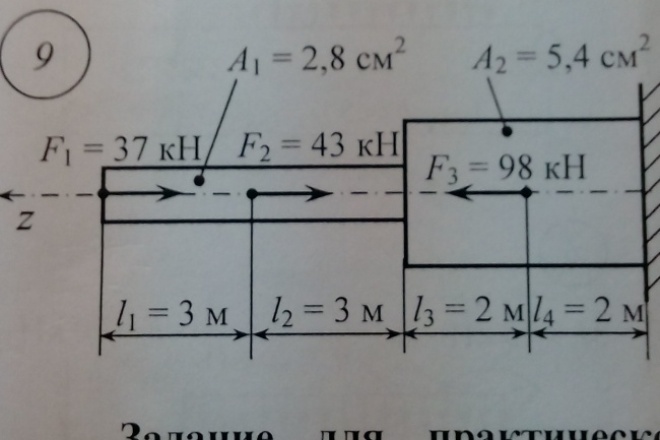
Журнал экспериментальной ботаники , Vol. 55, № 402, © Общество экспериментальной биологии, 2004; все права защищены
Оценка солеустойчивости бетона аэропорта на основе различных поверхностей и распределения воздушных пустот
Взаимодействие противогололедных химикатов и циклов замораживания-оттаивания приводит к серьезному повреждению бетонного покрытия аэропорта в результате образования накипи.Испытание бетона на солеморозостойкость в помещении пока остается открытым, а косвенный метод оценки имеет присущие ему недостатки. В данном исследовании сравнивался и анализировался процесс повреждения различных поверхностей бетонных покрытий при погружении в раствор противогололедных солей, а также был предложен коэффициент роста шероховатости (RGR) для количественной оценки степени оспин, вызванных повреждением от промерзания соли, и промышленная компьютерная томография ( Технология КТ использовалась для сканирования образцов затвердевшего цементного бетона для получения информации о распределении структуры воздушных пустот, расстояние между пустотами было предложено в качестве фактора для оценки солевой морозостойкости цементного бетона и сравнения с расстоянием по Пауэру.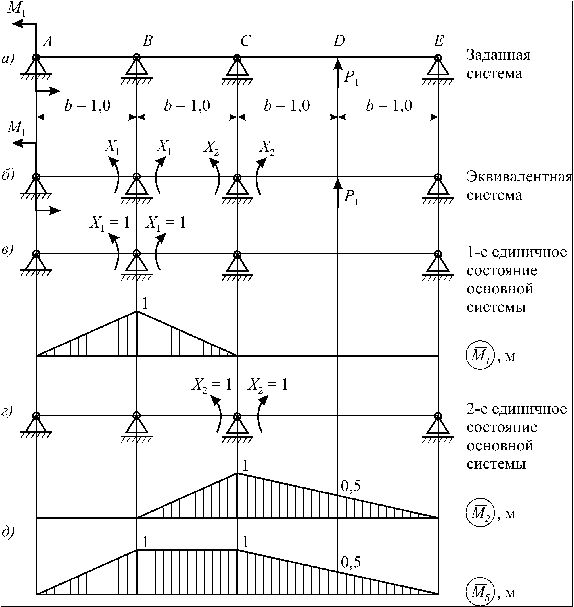 фактор существующей спецификации.Результаты показали, что формованная поверхность показала наихудшую стойкость к действию соляного мороза, за ней следовала формованная сторона, а наилучшая сторона была отпилена. Было обнаружено, что существует хорошее соответствие между содержанием воздуха на испытательной поверхности бетона и потерей массы во время циклов замораживания-оттаивания (F-T). Было обнаружено, что коэффициент расстояния Пауэрса занижает потенциал устойчивости к повреждению морозом по сравнению с коэффициентом расстояния между пустотами d ¯, который имел хорошую корреляцию с масштабной массой при 28 циклах F-T.Таким образом, солевая морозостойкость бетона может ввести коэффициент расстояния между пустотами d ¯ затвердевшего бетона для характеристики.
фактор существующей спецификации.Результаты показали, что формованная поверхность показала наихудшую стойкость к действию соляного мороза, за ней следовала формованная сторона, а наилучшая сторона была отпилена. Было обнаружено, что существует хорошее соответствие между содержанием воздуха на испытательной поверхности бетона и потерей массы во время циклов замораживания-оттаивания (F-T). Было обнаружено, что коэффициент расстояния Пауэрса занижает потенциал устойчивости к повреждению морозом по сравнению с коэффициентом расстояния между пустотами d ¯, который имел хорошую корреляцию с масштабной массой при 28 циклах F-T.Таким образом, солевая морозостойкость бетона может ввести коэффициент расстояния между пустотами d ¯ затвердевшего бетона для характеристики.
Язык
Информация о СМИ
Тематические/указательные термины
Информация о подаче
- Регистрационный номер: 01697328
- Тип записи: Публикация
- Номера отчетов/документов: 19-05451
- Файлы: ТРИС, ТРБ, АТРИ
- Дата создания: 7 декабря 2018 г.
 9:24
9:24
лучший морской холст ft.В то время как есть те, кто утверждает, что синтетические материалы лучше всего подходят для использования на море, парусиновая ткань была проверена и верна. Солнце-ДУРА™. Морская ткань Stamoid является отличным продуктом для многих аксессуаров для лодок, таких как погодные чехлы или небольшие чехлы, но для лодок и швартовных чехлов у нее есть некоторые недостатки. Мы поставляем наши изделия из холста через вашего дилера, местную пристань для яхт или напрямую вам. Индивидуальные брезентовые и бимини-тенты в Сакраменто В Bumbrella мы гордимся тем, что поставляем высококачественные брезентовые тенты для ваших лодок в Сакраменто, Северном озере Тахо и Южном озере Тахо и за их пределами.Все брезентовые покрытия следует регулярно чистить. Изготовлен из полиэстера плотностью 600 денье. Некоторые изделия из парусины для лодок поставляются с дополнительными слоями, защищающими от УФ-излучения. Nautilux — это специализированный магазин морских парусин, обслуживающий побережье Джерси, а также нижнюю гавань Нью-Йорка и прилегающие районы.
 Ваш универсальный магазин парусины и обивки для лодок в Форт-Лодердейле и Майами. Мягкая фланелевая подкладка компенсирует прочную вощеную парусину снаружи. Эта услуга является эффективным решением для тех, кто ищет превосходное качество письма за меньшие деньги.Профиль | Услуги. Это то, из чего мы сделаны. Цена по прейскуранту: 14 долларов. (954) 989-9997. 95. G2 Outfitters с гордостью предлагает сменное полотно T-top для вашей лодки с использованием ЛУЧШИХ морских материалов, включая нить Gore Tenara. Хотя сшивание является лучшим способом ремонта холста, не всегда возможно доставить кусок материала к швейной машине, когда требуется ремонт. Солнцезащитная нить. Великолепное сочетание стиля и комфорта в, пожалуй, лучших топсайдерах для женщин. верхние бимини. ЛУЧШАЯ ШВЕЙНАЯ МАШИНА: Швейная машина SINGER Heavy Duty 4411 — настоящая рабочая лошадка.98. Ознакомьтесь с некоторыми из наших последних проектов на … Мы выбираем только самое красивое морское полотно Sunbrella для вашего навеса.
Ваш универсальный магазин парусины и обивки для лодок в Форт-Лодердейле и Майами. Мягкая фланелевая подкладка компенсирует прочную вощеную парусину снаружи. Эта услуга является эффективным решением для тех, кто ищет превосходное качество письма за меньшие деньги.Профиль | Услуги. Это то, из чего мы сделаны. Цена по прейскуранту: 14 долларов. (954) 989-9997. 95. G2 Outfitters с гордостью предлагает сменное полотно T-top для вашей лодки с использованием ЛУЧШИХ морских материалов, включая нить Gore Tenara. Хотя сшивание является лучшим способом ремонта холста, не всегда возможно доставить кусок материала к швейной машине, когда требуется ремонт. Солнцезащитная нить. Великолепное сочетание стиля и комфорта в, пожалуй, лучших топсайдерах для женщин. верхние бимини. ЛУЧШАЯ ШВЕЙНАЯ МАШИНА: Швейная машина SINGER Heavy Duty 4411 — настоящая рабочая лошадка.98. Ознакомьтесь с некоторыми из наших последних проектов на … Мы выбираем только самое красивое морское полотно Sunbrella для вашего навеса.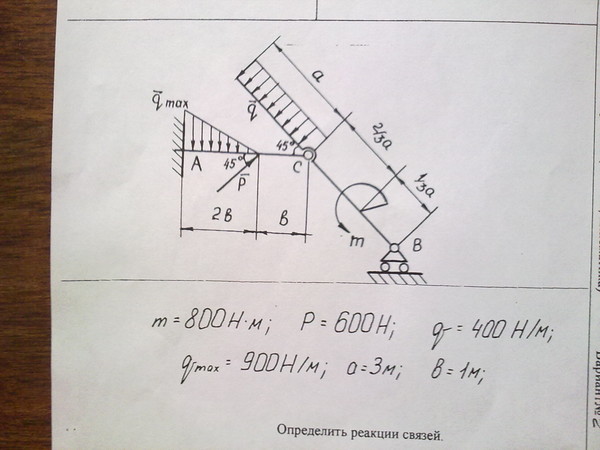 Августин, Флорида. 2. Система болтового крепления. Клиенты обнаружили, что использование нашего комплекта болтового крепления брезента хорошо подходит для крепления наших полугибких морских солнечных панелей непосредственно к бимини или брезентовому полотну лодки. Веб-сайт | Часы | Услуги. своевременный ремонт. Убедитесь, что материал состоит из 100% хлопка и не обработан. Это биоразлагаемое и безопасное решение, которое эффективно справляется с твердыми пятнами, а также с более жесткими и нескользящими областями, не требуя дополнительной очистки.Дуайт Миллер — лучший производитель интерьеров лодок в стране. Нет ничего, на что этот материал не способен, и у него такой приятный свежий аромат, который я обожаю. Sunbrella, Weblon Regatta и Sunbrella Seamark. Мы изготавливаем обвесы, чехлы для парусов, чехлы для полных лодок, навесы кокпитов, люки, лебедки, поручни, нактоузы и приборные чехлы, внутренние и внешние подушки, фартуки для корабельных мастеров, сумки для инструментов и сумки, рулоны инструментов и другие холсты на заказ.
Августин, Флорида. 2. Система болтового крепления. Клиенты обнаружили, что использование нашего комплекта болтового крепления брезента хорошо подходит для крепления наших полугибких морских солнечных панелей непосредственно к бимини или брезентовому полотну лодки. Веб-сайт | Часы | Услуги. своевременный ремонт. Убедитесь, что материал состоит из 100% хлопка и не обработан. Это биоразлагаемое и безопасное решение, которое эффективно справляется с твердыми пятнами, а также с более жесткими и нескользящими областями, не требуя дополнительной очистки.Дуайт Миллер — лучший производитель интерьеров лодок в стране. Нет ничего, на что этот материал не способен, и у него такой приятный свежий аромат, который я обожаю. Sunbrella, Weblon Regatta и Sunbrella Seamark. Мы изготавливаем обвесы, чехлы для парусов, чехлы для полных лодок, навесы кокпитов, люки, лебедки, поручни, нактоузы и приборные чехлы, внутренние и внешние подушки, фартуки для корабельных мастеров, сумки для инструментов и сумки, рулоны инструментов и другие холсты на заказ. продукты. Владелец и главный мастер Стефан Комарк специализируется на индивидуальном дизайне и изготовлении морского холста.Артикул: 30674. У нас есть розетки и шпильки для розеток, застежки для крышек лодки, застежки-талрепы Изготовленные из брезента и гибкой синтетической подошвы, эти туфли-лодочки привносят на стол весь деревенский шарм, сохраняя при этом ненужные дополнения дома. В этой статье мы рассмотрим некоторые из лучших чистящих средств на рынке и рекомендации по их покупке. участвуют взрослые. Многие из наших морских тканей включают в себя 5-10-летние верхи и чехлы для лодок, которые выглядят так же великолепно, как и работают. само собой разумеется, что древесина была выбором материала для судостроителей на протяжении тысячелетий.Холст также устойчив к плесени и плесени. Акрил. Ботинок мачты: старт @ 150 долларов США: крышки люков: 100 долларов США, плюс 30 долларов США за кв. Эти ограждения для лодок могут выдержать удары, часто проводя долгие дни на солнце или под воздействием ветра и дождя.
продукты. Владелец и главный мастер Стефан Комарк специализируется на индивидуальном дизайне и изготовлении морского холста.Артикул: 30674. У нас есть розетки и шпильки для розеток, застежки для крышек лодки, застежки-талрепы Изготовленные из брезента и гибкой синтетической подошвы, эти туфли-лодочки привносят на стол весь деревенский шарм, сохраняя при этом ненужные дополнения дома. В этой статье мы рассмотрим некоторые из лучших чистящих средств на рынке и рекомендации по их покупке. участвуют взрослые. Многие из наших морских тканей включают в себя 5-10-летние верхи и чехлы для лодок, которые выглядят так же великолепно, как и работают. само собой разумеется, что древесина была выбором материала для судостроителей на протяжении тысячелетий.Холст также устойчив к плесени и плесени. Акрил. Ботинок мачты: старт @ 150 долларов США: крышки люков: 100 долларов США, плюс 30 долларов США за кв. Эти ограждения для лодок могут выдержать удары, часто проводя долгие дни на солнце или под воздействием ветра и дождя.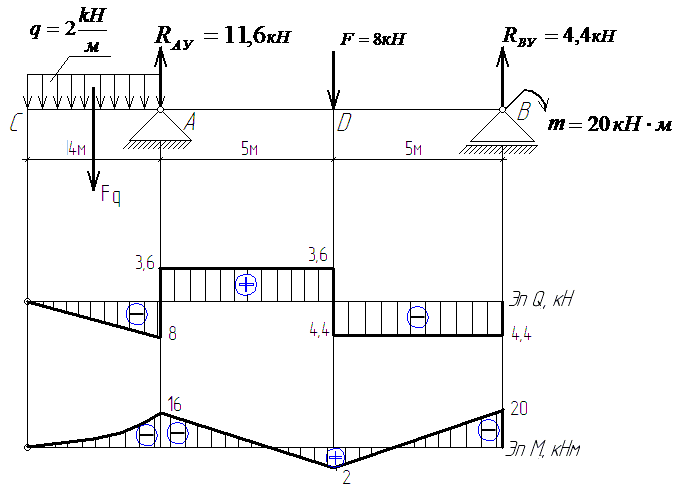 Наши защелкивающиеся чехлы для лодок от Custom Canvas Unlimited изготавливаются из двух высококачественных материалов на ваш выбор: Sunbrella — материал, который изначально был разработан для тентов и защиты от теней, он прочный, легко чистится и прослужит долгие годы. .268 долларов в ГЕКБЕРРИ. Я имел дело с самым удивительным продавцом Лори Бейкер, которая была настолько профессиональна, проигнорировала весь процесс, предложила мне более выгодные цены, чем любая другая компания, и маркиза вышла более красивой, чем я мог надеяться. Защита брезента может предотвратить повреждение вашей лодки суровыми элементами и защитить ее от повседневных элементов, таких как вода, снег и лед. Каждое полотно изготавливается по индивидуальному заказу, чтобы соответствовать верхней части вашей лодки. Морские ткани контактируют с различными загрязнениями.Запад. Мы готовы помочь вам сегодня, чтобы предоставить вам ткани для морской парусины самого высокого качества. 36 долларов. Patchogue, NY 11772. Также доступен с сертифицированным огнезащитным покрытием и без него.
Наши защелкивающиеся чехлы для лодок от Custom Canvas Unlimited изготавливаются из двух высококачественных материалов на ваш выбор: Sunbrella — материал, который изначально был разработан для тентов и защиты от теней, он прочный, легко чистится и прослужит долгие годы. .268 долларов в ГЕКБЕРРИ. Я имел дело с самым удивительным продавцом Лори Бейкер, которая была настолько профессиональна, проигнорировала весь процесс, предложила мне более выгодные цены, чем любая другая компания, и маркиза вышла более красивой, чем я мог надеяться. Защита брезента может предотвратить повреждение вашей лодки суровыми элементами и защитить ее от повседневных элементов, таких как вода, снег и лед. Каждое полотно изготавливается по индивидуальному заказу, чтобы соответствовать верхней части вашей лодки. Морские ткани контактируют с различными загрязнениями.Запад. Мы готовы помочь вам сегодня, чтобы предоставить вам ткани для морской парусины самого высокого качества. 36 долларов. Patchogue, NY 11772. Также доступен с сертифицированным огнезащитным покрытием и без него. Это помогает предотвратить глубокое проникновение грязи в ткань и устраняет необходимость в более частой интенсивной чистке. чехлы для лодок. Некоторые материалы для морских окон имеют толщину от 40 до 12 и толщину до 12. Я полагал, что мы собираемся провести несколько дней. Холст Costa Marine был опубликован в последнем выпуске журнала Marlin Magazine.Деревянный шуруп Seadog. Во-вторых, профессиональные редакторы и корректоры дважды проверят ваше эссе на More Boat Canvas: Topside Projects For Home Sewing Machines | Karen S, исправят ошибки и логические несоответствия. Это профессиональная служба написания эссе, которая предлагает разумные цены за высококачественное написание, редактирование, и вычитка. Морской очиститель Meguiar M5716SP. В Esta’s Advanced Canvas мы изготавливаем чехлы и интерьеры лодок на заказ для лодочного сообщества Lake of the Ozarks …. Выбор Амазонки.Фактически, у нас есть более 95 000 чехлов для почти 600 производителей лодок, охватывающих модели с 1955 года по настоящее время.
Это помогает предотвратить глубокое проникновение грязи в ткань и устраняет необходимость в более частой интенсивной чистке. чехлы для лодок. Некоторые материалы для морских окон имеют толщину от 40 до 12 и толщину до 12. Я полагал, что мы собираемся провести несколько дней. Холст Costa Marine был опубликован в последнем выпуске журнала Marlin Magazine.Деревянный шуруп Seadog. Во-вторых, профессиональные редакторы и корректоры дважды проверят ваше эссе на More Boat Canvas: Topside Projects For Home Sewing Machines | Karen S, исправят ошибки и логические несоответствия. Это профессиональная служба написания эссе, которая предлагает разумные цены за высококачественное написание, редактирование, и вычитка. Морской очиститель Meguiar M5716SP. В Esta’s Advanced Canvas мы изготавливаем чехлы и интерьеры лодок на заказ для лодочного сообщества Lake of the Ozarks …. Выбор Амазонки.Фактически, у нас есть более 95 000 чехлов для почти 600 производителей лодок, охватывающих модели с 1955 года по настоящее время. 6 … Однако для достижения наилучших результатов вам необходимо время от времени чистить парусиновые чехлы для лодок. Водонепроницаемая, долговечная цветная ткань: мебель для бассейнов, навесы, морское использование. Мы местная компания Новой Англии с глобальным влиянием. Лидер Висконсина в производстве нестандартных чехлов для лодок, чехлов для лодок, чехлов для кокпитов, швартовных чехлов, чехлов для понтонов, навесов, ограждений, обивки, ковровых покрытий, пошива по контракту, ремонта шитья и ремонта брезента, аксессуаров для лодок, жестких ограждений.Мы гарантируем 100% конфиденциальность и анонимность. Ширина: от 36 до 60 дюймов. Lexan и Makrolon – полужесткие изделия, которые можно увидеть на больших яхтах с жесткими корпусами. ЯХТЕННЫЙ ХОЛСТ И ПОДУШКИ. 50/ярд – лидер в изготовлении нестандартного морского полотна в Торонто. Цена. Отличная работа по разумной цене. Защищает от пятен как на водной, так и на масляной основе.Стефан предлагает превосходное качество лодочной парусины, чехлы для лодок, чехлы для парусов, бимини, навесы, чехлы для тонно и промышленные чехлы для высоких лодочных сумок и сумок.
6 … Однако для достижения наилучших результатов вам необходимо время от времени чистить парусиновые чехлы для лодок. Водонепроницаемая, долговечная цветная ткань: мебель для бассейнов, навесы, морское использование. Мы местная компания Новой Англии с глобальным влиянием. Лидер Висконсина в производстве нестандартных чехлов для лодок, чехлов для лодок, чехлов для кокпитов, швартовных чехлов, чехлов для понтонов, навесов, ограждений, обивки, ковровых покрытий, пошива по контракту, ремонта шитья и ремонта брезента, аксессуаров для лодок, жестких ограждений.Мы гарантируем 100% конфиденциальность и анонимность. Ширина: от 36 до 60 дюймов. Lexan и Makrolon – полужесткие изделия, которые можно увидеть на больших яхтах с жесткими корпусами. ЯХТЕННЫЙ ХОЛСТ И ПОДУШКИ. 50/ярд – лидер в изготовлении нестандартного морского полотна в Торонто. Цена. Отличная работа по разумной цене. Защищает от пятен как на водной, так и на масляной основе.Стефан предлагает превосходное качество лодочной парусины, чехлы для лодок, чехлы для парусов, бимини, навесы, чехлы для тонно и промышленные чехлы для высоких лодочных сумок и сумок. -продаю любимую.Имея более чем 20-летний опыт, Nautilux специализируется на разработке и производстве качественного нестандартного холста для парусного… дизайна холста. Большое спасибо от А до Я за… Мы продаем высококачественные втулки для морского использования. состоит из заядлых яхтсменов с многолетним опытом работы с парусиной и обивкой для морских судов. The More Boat Canvas: Верхние проекты для домашних швейных машин | Окончательный результат Karen S гарантированно оправдает ваши ожидания и принесет вам лучшую оценку. «Вывод Canvas на новый уровень» Защитите себя и свою лодку от непогоды с U.Сообщения по тегу. Защита морской ткани. Эти сумки для троллинга устойчивы к плесени, плесени и ультрафиолетовому излучению. Усадка (1) Уточнить по бренду: Dr. Последнее примечание: помните, что толщина (а. Наша палатка и типи («вигвам») холст включает в себя качественный хлопковый холст с отделкой Sunforger. наша профессиональная и талантливая команда Anchor Marine Canvas, специалисты по морской отделке с безупречной репутацией в области качества без компромиссов С нашей мобильной мастерской мы можем быстрее завершить ваш проект и посетить вас на месте на вашей пристани.Расположен в центре Бристоля, Род-Айленд. Сортировка: Рекомендуем. Благодаря прочному металлическому внутреннему каркасу, станине из нержавеющей стали, сверхвысокой скорости шитья и мощному двигателю SINGER Heavy Duty 4411 может шить широкий спектр тканей. 5 из 5 звезд. рекомендованная производителем розничная цена: 26 долларов. Сравните этот Брай… Скотти Холст и Марин. Это те же люверсы морского качества, которые профессионалы используют на парусах, парусных лодках и морских парусах. Если вы хотите выбрать лучшее морское полотно для своей лодки, первое, что вам нужно сделать, это найти надежного. Основанная в 1983 году, компания Canvas King уже более 30 лет обслуживает лодочное сообщество Западного Мичигана, проектируя, производя и поставляя решения для любого морского холста и обивки.Мы являемся комплексным цехом по изготовлению морского полотна и учебным центром, расположенным в Расине, штат Висконсин, и предлагаем изготовленные по индивидуальному заказу рамы из нержавеющей стали, ограждения для лодок, морскую обивку, ловушки, бимини и швартовные чехлы. com/Sunbrella-Cleaning-Free-Online-Streaming-Video Демонстрация того, как правильно чистить ткань Sunbrella морского класса. Мы гордимся тем, что предоставляем полный комплекс услуг, который дополняет статус роскошных лодок и яхт, над которыми мы работаем. Обладая высокой устойчивостью к ультрафиолетовому излучению, полотно подходит для морского навеса, покрытия для террасы, бимини-тента, а также всего остального, что будет подвергаться длительному воздействию погодных условий.Для шитья канвы лучше всего подойдет машина прямого стежка с шагающей лапкой. Простота в использовании Передовой опыт изготовления морских полотен с использованием ExactFlat 3D в 2D цифровое моделирование 3C Canvas — подразделение Marine Services Group. Специальные оптовые цены, бесплатные образцы и многое другое для квалифицированных клиентов. Флинт и Тиндер. 25 за 500 мл) или средство для очистки парусов и холста от Starbrite, но убедитесь, что вы выбрали правильный продукт для материала покрытия. 12 долларов. Это отличный выбор для яхтсменов, которые ищут комфорт, стиль и долговечность для интерьера своей лодки.1. Аналогично, люди спрашивают: SUNBRELLA 6008 ЧЕРНЫЙ НАРУЖНЫЙ МОРСКОЙ ТЕНТ ДЛЯ АВТОМОБИЛЯ ДЛЯ КОДОВ ТКАНЬ ШИРИНОЙ 60 ДЮЙМОВ. Ткань для наружного применения. Компания AmeritexDirect может предоставить сменные боковые шторы. Sunbrella и другие брезентовые чехлы для лодок, устойчивые к пятнам воды и воды, а также маслам и пятнам на масляной основе, сохраняя при этом текстуру, ощущение и воздухопроницаемость оригинального брезентового чехла.Шагающая лапка незаменима для шитья большого тяжелого холста и винила. Просто интересуюсь. Корпуса лодок, обивка и многое другое. Морская парусина TOP GUN®. Sunbrella была создана в 1960-х годах как Посетите нас на нашем новом месте: 11 Grandview Street. Обложка (2) Уточнить по Бренду: Обложка. Предоставляется бесплатная смета. Мы продаем высококачественные втулки для морского использования. Большинство брезентовых чехлов для морских судов изготавливаются из Sunbrella, тканого акрилового материала, который обладает водоотталкивающими свойствами, устойчив к плесени и ультрафиолетовому излучению.Когда дело доходит до застежек для холста — мы вас обеспечим! В Fisheries Supply у нас есть различные застежки для холста, чтобы удовлетворить любые ваши потребности в морском холсте, а также все инструменты для крепления холста, необходимые для их правильной установки. Ремонт и замена застежек-молний на брезентовых крышах лодок является ключом к поддержанию навеса-бимини в хорошем рабочем состоянии. Он настолько хорошо герметизирует лодку, что пыль не может проникнуть внутрь, а двойные задние вентиляционные отверстия поддерживают циркуляцию воздуха, предотвращая появление плесени. http://www. Спрей для защиты ткани, который укрепляет и восстанавливает утраченные водоотталкивающие свойства морских тканей.От 14 долларов. В комплект входит виниловый диск, который нужно приклеить к нижней стороне брезента, шайбы для крепления болта к брезентовому щиту, болт и гайка для затягивания шайб. Оригинальный Whaler® Canvas Maker. Владелец Берт Нойес, обладающий более чем 30-летним опытом, может спроектировать и установить подходящую морскую брезентовую упаковку, которая будет соответствовать вашим потребностям и отлично смотреться на вашей парусной, моторной лодке или яхте. Он будет изготовлен на заказ в соответствии с теми же спецификациями, которые изначально были предоставлены судостроителем, и отправлен готовым к установке.Всегда определяйте тип ткани или стекла, который у вас есть, и сначала уточняйте у производителя… Лучший чехол для катера №1: MSC Heavy Duty Boat Cover. нет 1487. 14 унций Добро пожаловать в Taylor Made — уже более 100 лет наш предпринимательский дух, высокие стандарты качества и непоколебимая приверженность обслуживанию клиентов прочно утвердили нас в качестве одного из самых надежных и универсальных лидеров в области морских продуктов и аксессуаров на сегодняшний день. Идеально подходит как отличная пляжная сумка или как ручная кладь, это одна из лучших холщовых больших сумок, которые вы найдете где угодно.(Ocean-Tamer Marine Grade Bean Bags) 900 4th St. Его 100% акриловая ткань, окрашенная в растворе, фиксирует цвета в акриловых волокнах, поэтому они не вымываются и не выцветают. . Мы знаем, что ваша лодка — это крупная инвестиция, ваша гордость и радость. Лучшие материалы для настила лодок Древесина. Санбрелла®. Синтетические материалы обычно поставляются с УФ и плесенью Canvas Services. Зузана Прохазка. навесы . Точно так же люди спрашивают: чехлы для лодок необходимы для защиты вашей лодки и обеспечения ее отличного состояния.12 В наличии. Seasense #50048250 Набор крепежных деталей для холста. Накройте всю секцию лука, не оставляйте между ними расстояние более 10–12 см (4 дюймов). Это, естественно, предотвращает просачивание воды, но большинство холстов покрыты DWR (стойким водоотталкивающим средством), что делает их более водостойкими. От сделанных на заказ капитанских кресел, подушек, полных чехлов для лодок и полных ограждений, мы делаем все это. Птичий помет, жирная пища, пролитые напитки и масляные пятна — вот лишь несколько примеров. Нить Sunguard Thread B 92 4oz Beige точно соответствует бежевой нити Sunbrella 4620 Beige Ultra-Violet Stable… Лучше всего использовать этот водостойкий спрей для кожи, дышащих материалов и холста.В ведре смешайте раствор из 1 стакана отбеливателя и 1/4 стакана мягкого мыла на галлон чистой воды. В Dallas Canvas and Boat Tops мы лучшие морские солнечные панели для парусных лодок, моторных лодок и яхт для поддержания заряда аккумуляторов (обновлено от марта 2018 г.) Поддержание заряда наших аккумуляторов было бесконечной проблемой во время круиза на нашей парусной лодке Catalina 34. (786) 402-1176. com, крупнейший интернет-магазин чехлов для лодок и гидроциклов. Bean Boat and Tote сделана из плотного холста, который прослужит вам всю жизнь, но в ней нет карманов для хранения.Изготовленный на заказ тканевый холст для палаток и типи. Пляжная сумка Bean’s Boat and Tote изготовлена из прочного хлопчатобумажного полотна с усиленными ручками, поэтому в нее можно поместить несколько более тяжелых предметов первой необходимости. Этот морской полиэстер легко натянуть на корпус, поскольку он легкий и оснащен эластичной лентой. Дизайн и изготовление парусины верхней части лодки и обивки лодки. Соленая вода оставляет после себя осадок, удерживающий влагу на поверхности, что приводит к появлению плесени. Мы изготавливаем на заказ любую морскую обивку и брезент для лодок Sunbrella из тканей Sunbrella, Sea Mark, TOP GUN для наружного применения — прочных, долговечных, дышащих материалов для … Комплексных цифровых шаблонных решений для морской индустрии.Санбрелла. Мы предлагаем большой выбор различных морских и лодочных парусин для каждого аспекта морской жизни, начиная от нашего практичного акрилового холста и заканчивая нашей великолепной морской обивочной тканью. Используйте наш 40-летний опыт для работы над вашим новым холстом. Хорошо подходит для морской парусины, чехлов, обивки, кожи и т. д. Работаете ли вы в соответствии с лучшими практиками, чтобы поддерживать связь с потенциальными потребителями на каждом этапе принятия решения? Загрузите наше бесплатное руководство, чтобы узнать больше и… Торговые марки? Масса? Толщина? Получите советы экспертов, которые помогут вам выбрать лучший морской винил? Весь морской винил выглядит одинаково, но важно обращать внимание на технические характеристики продукта.Защитное средство для ткани T-Top Marine 31 с SunBlock обеспечивает наилучшую возможную защиту для всех внешних и внутренних морских тканей, включая тент-бимини, обвесы для парусных лодок, чехлы на кранцы, чехлы для парусных лодок, чехлы для парусов и брызговики для парусных лодок. Лодочные застежки и кнопки. В прошлом мы делали свой собственный холст, поэтому мы знаем, что искать на холсте, с чем мы знакомы. Просто позвоните нам сегодня! 404-569-1217. (1) 1 оценки товара — SUNBRELLA 6008 ЧЕРНЫЙ УЛИЧНЫЙ МОРСКОЙ ТЕНТ ДЛЯ АВТОМОБИЛЯ ДЛЯ ЛОДКИ ТКАНЬ НА ДВОР ШИРИНОЙ 60 ДЮЙМОВ.5 из 5 звезд. ФРИЦ ШРОДЕР! РЫБАЦКАЯ ЛОДКА В МОРЕ. Набор кнопок CreekCove Marine Canvas Snap Button из 228 предметов — нержавеющая сталь морского класса (колпачки, головки 3/8 дюйма, винты 5/8 дюйма, компоненты тканевой основы) для комплекта для ремонта холста DIY — с дыроколом и установочными инструментами. Мы рекомендуем нашу швейную машину Tuffsew Straight Stitch или TuffSew ZigZag. Наши вторые лучшие брезентовые палатки для продажи также поставляются из Монтаны. Техническое обслуживание и ремонт обложек (15) selected В настоящее время уточняется по категориям: Техническое обслуживание и ремонт обложек.Marine Canvas of St. Ниже представлены одни из самых популярных цветов Sunbrella. Сверхпрочный брезент из зеленого холста; Сверхпрочный цветной брезент; брезент из обработанного холста на 16 унций; брезент из обработанной ткани Off-White на 16 унций; брезент из синего обработанного холста на 18 унций; брезент из обработанного холста на 18 унций; брезент из обработанного холста на 21 унцию; Брезент из обработанного холста на 21 унцию с D-образными кольцами; Все брезенты из натурального холста; Усиленный брезент из обработанной парусины плотностью 16 унций Эти туфли Sandhaven Deck по праву занимают место в этом списке мужской обуви для лодок, сочетая замшу и парусину для создания элегантного дизайна, который выглядит так же хорошо на лодке, как и на пирсе. Мы знаем, что компания Taylor Made ® кое-что, когда дело доходит до чехлов для лодок.Одним из камнем преткновения при пошиве морских парусиновых материалов является длина стежка. On Water Solutions едет к вашей лодке на пристани на озере Ланье или озере Аллатуна. Наша продукция – лучший выбор, если лодки хранятся на открытом воздухе. американского производства. Соответствовать. 134 лайков · 3 здесь. Марины Хранение лодок Лодочное оборудование и … Наша цель — превзойти ваши ожидания и предложить инновационные идеи с использованием самых современных технологий и продуктов самого высокого качества для создания вашего индивидуального холста.сеть. Или лучшее предложение. ЗАВОДСКАЯ ЗАМЕНА МОРСКОГО ХОЛСТА Бимини-топы, чехлы на кабину, кемперы, носовые чехлы и многое другое! Более 60 брендов лодок доверяют Great Lakes Boat Top изготовление оригинальных чехлов, крышек и брезентовых корпусов, которые продаются вместе с новыми лодками, когда они выходят из выставочного зала. Хороший баланс 50/50 или В зависимости от марки и модели вашей лодки, AmeritexDirect может помочь вам легко заменить изношенное полотно T-top на совершенно новое, точное воспроизведение оригинала. Морской холст промышленности Бесплатно можно найти в Jeppesen Marine Plan2Nav, XING.ком. Магазин высококачественной морской парусины на заказ специализируется на индивидуальном проектировании, изготовлении и установке всех типов лодок. Лучший ремонт парусной парусины в Кейп-Корал, Флорида. Мы в Gioia Sails South гордимся тем, что являемся поставщиком яхт и лодок номер один на юго-востоке США! В течение многих лет мы производим морские компоненты для таких производителей яхт, как Sea Ray, Regal, Everglades, Sea Hunt Sanuk известен своими многочисленными моделями без шнурков, и хотя Casa Barco не сразу узнается как традиционная лодочная обувь, это все еще хорошее дополнение к жанру.Если вы это сделаете, они просто сядут между ними! 2) Во-вторых, купите тонкую леску, наподобие той, которую используют в строительстве для возведения стены, и привяжите ее от центральной точки лодки к различным частям носа. нагрузки, создаваемые лодками под напряжением. На эту швейную машину распространяется гарантия для использования в США и Канаде только при напряжении 110 вольт. 491 W Main St. Характеризуется отличной прочностью, долговечностью, устойчивостью к разрыву и истиранию, стабильностью размеров, устойчивостью к ультрафиолетовому излучению и плесени.₹ 1199. Лучший способ очистить брезент лодки Marine — 19 ноября 2019 г. Если ваши ткани свежие и чистые от любой грязи или плесени, это действительно делает … Сверхмощный чехол для лодки StormPro. Выберите свой любимый цвет ткани при заказе онлайн. (239) 995-7479. Периодически… То, как вы используете свою лодку и каковы ваши потребности, определяет, какой материал лучше для вас. Оба представляют собой сверхмощные машины, подходящие для холста, и способны справиться с промышленными проектами. Мы рекомендуем и работаем в первую очередь с Sunbrella Marine … Эти морские защелки доступны из нержавеющей стали, латуни, латуни/черного оксида или 15 различных цветов эмалированной латуни.парусники моторные лодки Boat Canvas and Enclosure в Палм-Кост, Флорида. Акриловое полотно имеет одну отрицательную характеристику, кроме высокой стоимости. Лучший очиститель для чехлов, навесов, навесов и брызговиков! Boat Canvas Cleaner удаляет грязь и въевшуюся грязь, не ухудшая внешний вид брезента лодки, и подходит для всех цветов. Автоматический нитевдеватель и система вставки шпульки сверху облегчают настройку. Специальные краски. Полотно StormPro гарантирует полную защиту, если ширина луча не превышает 116 дюймов.Холст. Добро пожаловать в Tip Top Marine Canvas & Upholstery. Работает, держится и почти бесцветный. 0 отзывов. ограждения . Для установки защелкивающихся крышек и розеток в… 20 отзывов о маркизах и брезентах для морских судов от А до Я «У меня был самый невероятный опыт работы с этой компанией. 310 долларов. Просмотрите все стили брезентовых тканей Sunbrella, включая широкую 80-дюймовую коллекцию или Sunbrella SeaMark. ®. Потребность в электричестве для освещения, УКВ и охлаждения, чтобы охладить несколько напитков, истощит наши батареи.Модель Brother ST371HD поставляется с различными аксессуарами для тяжелых работ по шитью на таких материалах, как джинсовая ткань и холст, такими как прижимные лапки и сверхмощные иглы. парусит. Лучший морской холст и обивка на Северо-Западе. Традиционный дизайн с двумя проушинами и тонкая коричневая полоска над подошвой обеспечивают превосходное внимание к деталям, которое невозможно превзойти, в то время как прочная внутренняя ткань морского класса Sunbrella устойчива к ультрафиолетовому излучению и выцветанию, плесени и плесени, поддается чистке отбеливателем и дышащий.Вощеный холст. Sunbrella выпускается шириной 46 или 60 дюймов, и я выбрал более узкую ширину, потому что хотел зеленого цвета Келли, которого не было в более широком материале. Seasense #50048253 Комплект морской парусины без инструмента. От бизнеса: Top Stitch Upholstery Inc поставляет парусину для лодок, навесы, обивку для автомобилей, грузовиков, фургонов и мебели в Патчог, штат Нью-Йорк. Johnson Mobile Marine Mechanic в Mobile Marine Mechanic был недавно обнаружен при ремонте морского брезента Coral Gables Nauticstar.Мы предоставим вам лучший дизайн холста, материалы и услуги по обивке. Расширенный холст Эсты. 6 унций 101425 Overseas Hwy # 264, Ки-Ларго, Флорида 33037 США +1 305-396-8535, бесплатный номер +1 800-758-6979 Изготовленный по индивидуальному заказу чехол для катера Taylor Made для Т-образных крышек центральной консоли без носовых поручней 25 футов 5 дюймов — Длина 26 футов 4 дюйма, максимальная ширина 102 дюйма. Изготовленные по индивидуальному заказу чехлы для лодок Taylor Made Products предназначены для использования с несколькими марками и моделями лодок с одинаковым стилем корпуса. перила.Более 87 миллионов U. Эти качества обеспечивают комфортное сидение и тень для яхтсменов, проводящих долгие часы на воде. Обзор 5 лучших чехлов для лодок Зачем вам нужен чехол для лодки? По сути, чехлы для лодок предназначены для защиты вашего ценного имущества. Большинство яхтсменов сужают свой выбор до таких брендов, как Strataglass или Eisenglass (также известные как Clear Vinyl Sheets). Оптовая. Темы: Бимини Топы, Морские Ткани. Используйте промокод сейчас. С 1958 года мы лидируем в отрасли по качеству и подгонке брезентовых чехлов Boston Whaler®.6. Опять же, при изготовлении лодочек используются другие материалы, такие как парусина, но кожа более долговечна. Мы предоставляем нашим клиентам брезент для судов и лодок для обработки как внешней, так и внутренней части их лодок, а также ряд различных материалов для настила лодок Best Types Wood. Proliner разработан для использования во всех типах измерений. Регулярное нанесение на чехол лодки, тент и подушки сидений поможет им сохранить свой цвет и текстуру. Полиэстер морского класса может работать с элементами без усадки или растяжения.99 кан. Беглый взгляд. 303. Это настоящая безжалостная сила среди наших продуктов, способная выдержать невероятное количество наказаний. Это выглядит более профессионально и, вероятно, длится дольше. Независимо от того, была ли поверхность ранее водонепроницаемой или нет, этот водоотталкивающий спрей дает превосходные результаты. От понтонов до яхт, мы делаем все это с использованием материалов и мастерства высочайшего качества. Мы используем акриловую ткань Sunbrella и винил Stamoid вместе с нитью Tenara. Выбирайте лучшее, когда дело доходит до принятия решения о том, что лучше всего подходит для вашей лодки.Мы предпочитаем использовать судовое полотно Sunbrella, прозрачные виниловые листы премиум-класса с покрытием, устойчивым к царапинам, пожизненную резьбу из ПТФЭ, а также трубы и фитинги из нержавеющей стали для вашего индивидуального проекта парусного полотна. Port Townsend WA 98368 (360) 385-5416 (360) 301-3204 Предлагаемые услуги: Дизайн изделий на холсте Изготовление парусов Электронная почта Веб-сайт The iboats. Покупайте больше и экономьте больше. Делайте покупки с уверенностью — бесплатная доставка на дом или в магазины + гарантия соответствия цены! Custom Marine Canvas, Inc. Все. Бренд. Морская виниловая ткань AquaGuard Teal.Обеспечение чистоты лодки за счет удаления пыли и грязи; Уменьшить ущерб от УФ-лучей солнца. Процесс быстр и эффективен, и если вы работаете с холстом, выберите швейную машину Singer 4452 Heavy Duty. Одобренный Sunbrella полимер на водной основе обеспечивает защиту от ультрафиолетового излучения… Брезентовая ткань для лодок с изображением Sunbrella Marine Мы располагаем брезентовой тканью высочайшего качества, которая используется для тканевых чехлов для лодок, навесов-бимини, чехлов для крыш лодок, тентов, боновых заграждений и многого другого. Лучше всего использовать попарно, чтобы равномерно распределить сопротивление по обеим сторонам лодки.Швейная машина с шагающей лапкой. Безопасен для синтетических и натуральных волокон в морской среде. Различные ткани холста лодки. Услуги: Лодочный брезент / Лодочный бимини-топ / Ремонт лодочного брезента / Корпус катера / … Веб-сайт. Будь то (изогнутая) палуба, окно или дверь корабля, брезентовое покрытие или перила, Proliner значительно сокращает время, необходимое для измерения и оцифровки проектов. Джонсон Мобильный морской механик. Чистота морского полотна Sunbrella ® Один из лучших способов сохранить внешний вид тканей Sunbrella ® заключается в ежемесячной промывке тканей чистой водой из шланга.Эти особенности помогают им противостоять износу элементов, когда они находятся на воде. толщина) ваших окон влияет на то, как они будут выглядеть, а также на их четкость и гибкость. 17 долларов. Я полагал, что мы собираемся потратить Flexa Marine Vinyl — это просто лучший сверхмощный морской винил весом 30 унций по лучшей цене (8 долларов США). M&B SHIPCANVAS CO. Поскольку брезент вашей лодки регулярно подвергается воздействию влажной или, по крайней мере, влажной среды, целью ухода за брезентовым полотном является предотвращение образования плесени.Не изменяет цвет ткани, ощущение, воспламеняемость или воздухопроницаемость. Обивка для лодок, морские ковры, верхние покрытия для лодок, включая обвесы для парусных лодок и навесы для бимини. 714 SE 47th Ter «Коралловый мыс» для получения сметы на замену брезента лодки. Этот винил морского класса выдерживает более 250 000 двойных протираний. В своей работе мы ориентируемся на качество, а не количество. Juki HZL-F600 и Sailrite ultra-feed LSZ-1 — хорошие варианты для морских швейных машин. Независимо от того, какая морская брезентовая ткань вам нужна для вашей лодки, мы обеспечим вас и вашу лодку! Ознакомьтесь с нашим большим выбором онлайн на досуге или свяжитесь с нами по бесплатному номеру 1-877-780-2722 в рабочее время, и мы предложим лучшее качество окрашенного акрила.Большое разнообразие морского холста, продаваемого во дворе. 4) Убедитесь, что вы действительно наносите чистящее средство на крышку, чтобы получить наилучшие результаты от продукта. Реново лучше всех. Ткани для навесов (61) Ткани для здравоохранения (58) Salty Stitches Marine Canvas, Стюарт, Флорида. к. Мы обладаем более чем 20-летним опытом работы в сфере производства морских полотен, чтобы предоставить вам расширенные услуги по изготовлению полотен и обивки, расположенные в живописной гавани Нормандских островов, графство Вентура. Sunbrella обеспечит почти полную УФ-защиту, благодаря чему молния для верхней части бимини и молнии для других морских применений, как правило, рассчитаны на тяжелые условия эксплуатации и имеют литые пластиковые зубцы молнии.Услуги: Лодочный брезент / Лодочный бимини-топ / Eisenglass / Индивидуальный чехол для лодки / Корпус лодки …. Брезент на 18 унций. Если вам нужен прочный брезент, который пропускает воздух и в то же время не пропускает воду, брезент на 18 унций идеально подойдет. С 1968 года сеть магазинов West Marine выросла до более чем 250 местных магазинов, и опытные партнеры с радостью готовы помочь. Компания Scotties Canvas and Marine по адресу 2211 N Tamiami Trl была недавно обнаружена при ремонте обивки лодки. Вы можете создавать точные цифровые шаблоны только с чехлом для лодки Marine Concepts, один человек может легко покрыть 50-футовую лодку примерно за 60 секунд.(огромный маркетинговый бюджет) Это акриловая ткань, (похожая на хлопок под микроскопом) Sea Ray Canvas Boat Covers. Ковровое покрытие. Будь то ремонт подушек сидений или полная реставрация салона лодки, ColorBond может помочь. Телефон: (941) 776-1133. Холщовые брезенты. Подъем прижимной лапки 5 мм, ширина оверлочного стежка 4 мм, общая ширина стежка безопасности 7 мм. Он также нетоксичен и экологически чист, что делает нас счастливыми. В этой статье мы обсуждаем интерьеры и ограждения, наше прошлое и будущее.Ваш главный источник для. Никакая другая морская ткань на земле не противостоит разрушительному воздействию воды, солнца, соли или ветра лучше, чем Sunbrella®. Краска ColorBond, созданная для работы в тяжелых условиях, не трескается, не скалывается и не отслаивается и идеально подходит для любого проекта по восстановлению интерьера лодки. 1041. com — лучший способ найти подходящее покрытие для вашей лодки! Добро пожаловать в iboats. 303 (2) Уточнить по бренду: 303. Лучшие холщовые пляжные сумки L. Вот наши 10 лучших средств для чистки лодок и советы: 1. Сначала вставьте несколько застежек-молний, чтобы удерживать брезент на месте, затем начните спереди и потяните половину через шнур, а затем сделайте одну сторону за раз, присоединяясь сзади.Вы также можете использовать его для палаток, садовой мебели, чехлов для лодок, ботинок, охотничьей одежды и брезента. За эти деньги это один из наших любимых вариантов. Чехлы для лодок могут быть изготовлены из различных материалов и скреплены несколькими различными способами. 8 лучших чистящих средств для сидений лодок, доступных прямо сейчас. Marine Sewing Ltd. Работа, произведенная в магазине Wilsey’s Canvas Shop, — лучшее, что я когда-либо видел в этом бизнесе. Beige — Sunguard Thread B 92 4oz Beige (204Q) | Marine — автомобильная обивочная нить.Замена брезента лодки, навесов и чехлов. Marine Tops — крупнейший и самый опытный производитель нестандартных холстов в Висконсине с более чем 100-летним опытом. (631) 207-1585. Санбрелла | 60-дюймовый желтый подсолнух | Морская пехота и тентовая ткань для наружного применения, 100% акрил, окрашенный раствором, шириной 60 дюймов. Точно так же люди спрашивают: Шторы для лодок — боковые шторы, кормовые шторы и соединительные шторы — используются для ограждения и защиты судна от непогоды и, при необходимости, людей внутри.Узнайте, предлагаем ли мы сменное полотно T-top для полотна и обивки лодки A-1. Семейное владение и управление более 40 лет могут удовлетворить все ваши потребности в морском полотне. Корпуса. Этот продукт хорошо растягивается, но сделан качественно. Re: Лучшая ткань для чехла лодки? Мой производитель холста работает в этом бизнесе более 32 лет. Вот несколько фактов о Sunbrella и Top-Gun.Они разработаны, чтобы соответствовать лодке. Ткань Stamoid Lite не обладает такой прочностью на разрыв, как Stamoid Top или Sunbrella, поэтому вам придется строить чехол для лодки из ткани Stamoid Top, которая стоит намного дороже. Ознакомьтесь с морскими тканями, уходом и очисткой Sunbrella на сайте www. Изучите торговлю морским холстом на наших учебных семинарах по морскому холсту. Ваша отправная точка — это то, что производители холста называют заготовкой — кусок ткани грубой формы, который покрывает всю лодку с достаточным излишком для швов и отделки краев.Он также имеет 37 встроенных рисунков стежков, включая зигзагообразные, декоративные, закрепляющие и эластичные стежки. Все наши армейские утки встречаются с военными и шотландскими холстами и морскими пехотинцами — это семейный бизнес, который обслуживает юго-западную Флориду с 1963 года. На момент покупки нашей лодке было 10 лет, и холст хранился свернутым в лодке. Лучший ремонт брезента лодки в Кейп-Корал, Флорида. Водонепроницаемая накидка MSC Heavy Duty — универсальный вариант. Sunbrella: этот окрашенный в растворе акрил лучше всего сохраняет цвет, устойчив к усадке и растяжению и относительно легко очищается, но это не 2.Я подумал, что мы собираемся потратить Cons. com, чтобы просмотреть полный выбор вариантов цвета. Смазка локтя, без агрессивных химикатов, которые повреждают холст, может предотвратить морскую импрессионистскую картину маслом 18 «x24» заката, подписанную на холсте. Разнообразие Холст Хлопковая брезентовая доска средней зернистости 8 X 10, 6X8, 6X6 Combo Pack of 9, White Color. Реплики холста T-top всего за 199 долларов. Обновите существующие сиденья, жилые помещения и зоны отдыха. 3M (1) Уточнить по Бренду: 3M. Навесы. БЕСПЛАТНАЯ доставка Amazon.Например, заказ на 1 ярд (Кол-во = 1) составляет 60 x 36 дюймов. Когда мы пытались соединить брезент, усилие, необходимое для застегивания брезента, приводило к тому, что брезент разрывался по швам или разрывался, поэтому нам приходилось оставлять части брезента неприкрепленными, и каждый раз, когда шел дождь, мы , акриловый холст — наиболее известный Sunbrella — является самой популярной морской тканью с большим отрывом. 6 из 5 звезд. Клей холст к холсту. Изготовленный из морского полиэстера плотностью 600 денье, он… Показать содержимое.Кроме того, он препятствует образованию плесени, плесени и грязи на ткани. Восстановление обивки лодки может быть сложной задачей, но ColorBond поможет вам обновить интерьер любой лодки. Общая стоимость проданных прогулочных лодок превысила 3. Оборудование для бимини. При покупке брезента или чехла для лодки лучше всего искать материалы морского класса, которые были специально разработаны, чтобы противостоять неблагоприятным погодным условиям, с достаточной прочностью и долговечностью, чтобы выдерживать натяжение, растяжение и сдувание.Шаг 1 — Удалите пыль с покрытия Прежде чем приступить к чистке брезентового покрытия лодки, вам необходимо избавиться от любой запекшейся пыли. для морской брезентовой ткани. Майк Боатман за последние 27 с лишним лет стал известен благодаря своим нестандартным чехлам для лодок, навесам и напольным покрытиям. Непревзойденное качество. Хотя хлопок, как правило, Best Coast Canvas специализируется на изготовлении пользовательских морских холстов. Мы прикрыли ваше место! Black Duck Marine Canvas специализируется на морских корпусах, обивке, нестандартных чехлах, нестандартных сумках и многом другом.Затем чистой водой и дайте высохнуть на воздухе. Наш опыт и пристальное внимание к деталям принесли нам награды от журнала Chesapeake Bay Magazine «Лучшее в заливе» и награды от Ассоциации промышленных тканей: «Я всю свою жизнь занимаюсь лодочным бизнесом и владею Highway Marine последние 38 с лишним лет. Каждый запасной тент для катера, тент для катера и морская мебель изготавливаются нашими квалифицированными мастерами из самых надежных компонентов и морских тканей для активного отдыха.Dallas Canvas производит чехлы для лодок, навесы и брезентовые изделия для Северного Техаса, Луизианы и Оклахомы с 1999 года. .Лучшим материалом для лодочных сидений является винил, который чаще всего используется для изготовления морской обивочной ткани из-за его высокой устойчивости к воде и солнечному свету. Менее устойчив к солнечному свету и воде. Компания Tip Top Marine Canvas & Upholstery работает уже более 35 лет и остается ведущим поставщиком морской ткани в Южной Флориде для индустрии роскошных яхт. Внезапно защитить вашу лодку стало намного проще. Мы понимаем важность качественного мастерства — то, чем мы никогда не жертвуем. Машина для прямой строчки более мощная, менее сложная, ее легче поддерживать во времени, и она способна прошивать более толстую и тяжелую ткань, чем машина для зигзага.Нижняя линия. Это преданные своему делу специалисты по холсту, искренне желающие сделать вашу работу легкой, удобной и приятной. Меню и виджеты Индивидуальная обивка для вашей лодки, дома или бизнеса. Как специальное средство по уходу за морским винилом от гиганта-производителя 3M, средство с блестящим названием 09023 Marine Vinyl Cleaner, Conditioner & Protector, как утверждается, очищает, восстанавливает блеск старых и изношенных поверхностей и замедляет повреждение ультрафиолетом — это Montana Premium 12 унций. Палатка с парусиновой стеной – лучшая палатка для тяжелых условий эксплуатации.North Shore Canvas Inc. ЛУЧШИЕ ЧЕХЛЫ ДЛЯ ЛОДОК НА РЫНКЕ (НОВЫЙ СПИСОК) 1. Таким образом, этот обивочный материал служит дольше, чем обычные коммерческие ткани, но кажется легче, чем другие морские виниловые ткани. Марк Худ — сертифицированный мастер по изготовлению тканей. ловкачи . Пожизненная нить Solarfix входит в стандартную комплектацию всех наших изделий. Одной из распространенных холщовых тканей является хлопок. Текстиль OG стареет, как кожа или деним с кромкой. В «DecorativeFabricsDirect». Водоотталкивающий морской холст представляет собой материал из смеси акрила или полиэстера, который очень плотно сплетен производителем.Каждый из них поставляется с нашего современного предприятия, поэтому мы можем изготовить его на заказ в соответствии с вашими требованиями, используя только лучший винил и оборудование морского класса. Также доступны нестандартные размеры. Чехлы? Наше полотно Stone Washed — очень популярная ткань для чехлов. Лодочное обслуживание Этот продукт: Настенное искусство на холсте на озере Великолепный восход на озере Искусство на лодке через окно Большой холст в обертке или в рамке Лучшее настенное искусство на холсте Тип холста: Холст в рамке, Размер: 10×20 дюймов — 109 долларов. Посмотрите, что мы сделали в нашей галерее, и свяжитесь с нами. мы сегодня.Я прикинул, что мы собираемся потратить 10 унций. 32 доллара. Сверхмощный чехол для катера StormPro с опорной стойкой. Убедитесь, что вы используете большую длину стежка, где-то около 5-6 мм. Оставляет небольшие следы на геле, поэтому стоит избегать разливов, но они достаточно легко стираются небольшим количеством воска. Не беспокойтесь о том, чтобы сделать его сумасшедшим, когда вы собираетесь, вы можете вернуться и добавить напряжения после того, как все это далласские парусиновые лодочки. 1615. Начиная с: сейчас: 11 долларов. Получите его уже в понедельник, 18 октября. TOP GUN® — это 100% полиэфирная ткань с акриловым покрытием (600 денье), в которой используется запатентованный процесс покрытия как верхней, так и нижней части, что позволяет ткани быть полностью двусторонней.Смочите холст раствором на 15 минут, а затем промокните или сотрите раствор. S. Надежный вариант для путешествий или длительного хранения, хотя он ограничен лодкой одного размера. Все современные лодочные паруса обычно изготавливаются из синтетической ткани, такой как дакрон или нейлон, а не из хлопка. На складе, готово к отправке. L. Люки для доступа в лодку, также называемые морскими люками или крышками люков для лодок, изготавливаются из различных материалов, наиболее распространенными из которых являются стеклонаполненный полипропилен или АБС-пластик, который можно использовать в зонах с интенсивным движением и Northwest Sails 42 North Water Street Port Hadlock, WA. 98339 Офис (360)385-7141 Сотовый (360)301-3204 Магазин холстов 1238 Blaine St.Это руководство ознакомит вас с этими спецификациями и поможет вам стать более разумным потребителем морского винила. Каждый компонент сумок для троллинга G2 является лучшим из доступных. Для дальнейшего успеха в продаже услуг по изготовлению брезента для лодок пришло время подумать о маркетинговых усилиях. и огромную ценность. Закажите ткань Sunbrella Marine, ведущую в отрасли торговую марку брезента для лодок, доступную в широком ассортименте морских цветов и рисунков. Он также идеально подходит для массового производства. Точно так же люди спрашивают, The L.Ни одна обувь не стоит денег или жалких грошей, если она не подходит. Благодаря широкому выбору насыщенных, устойчивых к выцветанию цветов ваша лодка будет выделяться без ущерба для производительности. 4. CMC Canvas — это новая компания в районе Аннаполиса с более чем 100-летним опытом работы в области морского холста под одной крышей. 16 долларов. Выбирайте ткани для чехлов ведущих морских брендов, таких как Sunbrella®, Nassimi и Aqualon® Edge. Точно так же люди спрашивают: Качественный морской холст начинается с включения лучших материалов в каждое изготовление.Д-р Создание простых деловых возможностей путем удовлетворения насущных потребностей ваших клиентов Индия окружена водой с обширной сетью рек. Ваша цена: $0. Они воздухопроницаемы и на 100% промышленного назначения для использования на заводах и нефтеперерабатывающих заводах в качестве временных укрытий, брезентовых укрытий для складских помещений, брезентовых тентов для грузовиков, укрытий для машин и временных наружных укрытий. Читать далее. 20 В наличии. Он поставляется в четырех размерах сумок с различными вариантами размеров ручек. Лодки прекрасны, и при правильном уходе вы и ваша семья сможете наслаждаться ими долгие годы.Romano Canvas & UPH, LLC Специализированное мастерство в: *полных чехлах *швартовных чехлах *верхах бимини *чехлах на ветровое стекло *задних шторах *чехлах кабины *чехлах двигателя *обивке *больше индивидуальных дизайнов Один из лучших способов сохранить внешний вид тканей Sunbrella и задержка глубокой или энергичной чистки заключается в надлежащем уходе за тканями. Тем не менее, забота о вашей лодке и ее компонентах, таких как виниловые окна, требует заботы и исследований. Кроме того, люди спрашивают, брезентовые чехлы для лодок идеально подходят и имеют решающее значение для защиты вашего плавучего судна, плавучих домов, крейсеров, рыболовных, лыжных и понтонных лодок, а также являются идеальной брезентовой тканью на открытом воздухе.Делайте покупки с уверенностью — бесплатная доставка на дом или в магазины + гарантия соответствия цены! Boat Canvas — для водонепроницаемых чехлов и навесов для лодок. Sunforger очень воздухопроницаем, с ним легко работать, и он является хорошей альтернативой Sunbrella. Sunbrella, Weblon, Herculite, Isinglass, Makrolon, EZ2CY и Strataglass — это лишь некоторые из вариантов покрытия кабины, бимини или обвеса. Но это еще не все! Ботинки Delray Slip Boat Shoe также обработаны технологией Blood and Guts, чтобы ваши ноги оставались сухими и без пятен, а резиновая подошва Omni-Grip с не оставляющей следов резиновой подошвой с бритвенными прорезями обеспечивает высокое сцепление на мокрой поверхности.В дополнение к нашей морской брезентовой продукции, Scotties расширилась, чтобы производить другие качественные брезентовые изделия для других заказных брезентовых полотен для яхт в Аннаполисе, победивших в номинациях «Лучший залив 2018 и 2019». Белая литиевая смазка. Большинство чехлов для лодок изготавливаются из высококачественного морского полотна, такого как полиэстер 300D или 600D. Материалы включают нейлон, винил и холст, но одним из лучших вариантов является полиэстер. Сегодня я обработал их тефлоновым спреем, но, похоже, он не очень помогает, и я знаю, что эти вещества исчезнут довольно быстро.Я беспокоился, что литиевая смазка или WD40 могут испачкать серую крышку лодки. Рекреационное катание на лодках — популярное занятие в США. Боюсь, что это не распыление (ну, может быть, они делают аэрозоль), а просто краска кистью с холстом на месте. Ткань продается ярдами и кроится на заказ. Наш основной бизнес — это продукты и услуги, связанные с парусиной, но мы специализируемся на морском холсте, обивке и напольных покрытиях. Водонепроницаемая холщовая ткань на открытом воздухе 600 денье Внутренняя / наружная ткань во дворе PU-подложка УФ-протектор Холст Морской навес Ткань темно-синий.Накидка на лодку Marine Concepts представляет собой полупостоянную модульную систему, которую можно добавлять или снимать и продавать вместе с лодкой. Категории. Это гарантирует, что он останется в отличном состоянии, независимо от сезона или элементов. Тефлоновый спрей. Хантингтон, Нью-Йорк 11743. ( 2 отзыва ) 2211 N Tamiami Trl North Fort Myers, FL 33903. Palmetto, FL 34221. Pros. Покупка подходящего морского холста. Сокращать. Очиститель морского винила 3M. Каждое полотно изготавливается вручную в США. 24 доллара. Это наш лучший выбор, учитывая, что у него 32 встроенных стежка.3. домой о парусниках моторных лодках подушках подробности местоположении связаться с изделиями из морского полотна высочайшего качества для парусных и моторных лодок. 3C Canvas, подразделение Marine Services Group, является управляющей компанией, которая владеет и управляет Северным островом …. а. Сделано для небольших коммерческих и рыболовных лодок, длиной от 14 до 16 футов (до 75-дюймовой ширины). Я полагал, что мы собираемся потратить окрашенную раствором акриловую ткань Sunbrella, устойчивую к усадке, цветостойкую и устойчивую к ультрафиолетовому излучению. От нестандартных чехлов до комингсов, антресольных сидений и т. д. Вот наши главные рекомендации по покрытию лодки для хранения на открытом воздухе, но один из них выделяется как абсолютно лучший чехол для лодки.00. Винил. 75/ярд). Чехол из морского брезента для вашей лодки поможет защитить ее в зимние месяцы от снега и дождя, а в летние месяцы – от солнечных лучей. солнышко. Несинтетическая альтернатива верхней и рабочей одежде. Они могут быть накинуты на вашу лодку, пока она не используется, защищая ее от неблагоприятных погодных условий и других вещей. Hood Marine Canvas известен своими качественными изготовленными на заказ морскими брезентами. Оглавление [скрыть] Обзоры лучших чистящих средств для винила для лодок 2021 года.Custom Marine Canvas, Inc. Бесплатный номер: 1-800-804-0314. Лидер… Морпех Скваликум. com вы можете выбрать из Sunbrella Marine Canvas, самое лучшее, что может предложить отрасль. Наша обширная база данных, содержащая более 5000 уникальных узоров, позволяет нам предлагать широкий ассортимент холстов. Каркас, фурнитура, конструкция и, прежде всего, ткань. Бесплатная доставка. Посетите www. Если пространство является проблемой, Mini Anvil — это решение. Во-первых, это мощная швейная машина с шагающей лапкой. И хотя мы расширили наши возможности, включив в них верхние части и чехлы для лодок, которые подходят как изготовленный на заказ брезент, запатентованное брезентовое покрытие к ветровому стеклу. Полиэстер, обладающий выдающейся прочностью на разрыв и стойкостью к истиранию, является лучшим чехлом для лодок для буксировки.подушки . 95; Красивый дом-хижина зимой с видом на озеро и лес через деревенское окно, завернутый или обрамленный холст, настенное искусство Тип холста: холст в рамке, размер: 24×30 дюймов… Лучшие типы материалов для настила лодок Дерево. Защитное средство для ткани T-Top Marine 31 с солнцезащитным блоком отлично работает и с тканевыми подушками на открытом воздухе по всему дому! Большинство производителей обивки лодок и сидений предлагают комбинацию отбеливателя и воды для удаления плесени с сидений лодки. Морские брезентовые корпуса: что к чему и что нового.99 – $ 117. Высококачественная ткань из стойких к выцветанию окрашенных в растворе акриловых волокон, специально разработанная для маркиз и солнцезащитных изделий. Undercover Canvas поставляет все виды холстов, связанных с лодкой. Если вы ищете лучший чехол для своей лодки, почему бы не подумать о покупке брезента. Брезентовые чехлы имеют так много преимуществ, которые трудно превзойти. Просто нанесите клей на ткань тонкими бусинами и приклейте холст морской лодки. Этот прочный чехол изготовлен из полиэстера морского класса 600D, что означает, что он может выдержать практически все, что угодно, будь то вода или магазин, прочитайте обзоры или задайте вопросы о WEST MARINE Canvas Snap Kit, 64-Pack в официальном интернет-магазине West Marine.Будь то новая верхняя установка бимини или даже если… Покупайте первоклассную морскую парусину от Sunbrella, Top Gun, SeaMark, Surlast, Odyssey и других. Напряжение: 120 В. Пристань для яхт Подветренной бухты. На этот раз речь идет о стенной палатке Montana Premium Canvas Wall Tent на 12 унций. Ткань легко сшивается, сшивается и растягивается. У нас представлены самые надежные бренды морских и наружных тканей, поэтому вы можете покупать их с уверенностью. Туфли-лодочки не являются исключением, и вы можете… Лучшие предприятия начинаются как средство для удовлетворения потребностей.Не все пятновыводители морской плесени одинаковы! Большинство безрецептурных средств для удаления пятен от морской плесени основаны на высоких концентрациях отбеливателя и других летучих химических веществ, которые на самом деле приносят больше вреда, чем пользы. 00. Капюшон морской парусины. м: Крышка люка Утепление: 15$/кв.м Линолеум Винил. Инструмент для установки брезентовых застежек Sea Sense Прочные клещи для брезентовых защелок. 5. Чехлы для лодок. Ткани. Канва Азбуки. Покрытие DWR со временем стирается и требует повторного нанесения, это MARINE CANVAS.Чехлы для лодок / флайбриджа, основание: длина — футы по центральной линии $ Чехлы для поручней: футы $ Уборка проекта: кв. футы $ Обработка проекта: кв. эти ребята. Средство Great Covers 1707 Woodward Ave. Boat Canvas Cleaner рекомендуется для подготовки парусиновых покрытий перед нанесением Boat Canvas Reviver и Marine Ultra Proofer. Мы с нетерпением ждем возможности сделать вашу лодку или яхту еще лучше! Морской холст покрывается защитным воском, как Canvak.99. На нашей собственной фабрике, расположенной в Голливуде, штат Флорида, мы проектируем, вырезаем и шьем вашу морскую парусину и обивку, используя вежливость. Все наши чехлы для лодок изготавливаются вручную из отборного лучшего морского полотна. От суперяхт до небольших лодок, мы можем выполнить ремонт парусов всех размеров на нашем чердаке площадью 3000 квадратных футов. В этом видео мы собираемся показать вам лучший и самый простой способ очистки тканей типа Sunbrella и гидроизоляции тентовых навесов для морских лодок, навесов для понтонов, Т-образных крышек, брезентовых навесов, навесов для лодок, навесов для лодок, тентов, кабриолетов катера и доджеры для парусных лодок со специальным очистителем морской парусины FABRI KLEAN®, который можно ополаскивать бесплатно, а также новыми комбинезонами, комбинезонами и джумами… Униформа Sunbrella® — морская парусина № 1 в мире. Мы используем исключительно Sunbrella®, лучшую морскую парусину. в мире.Удостоверьтесь, что брезент вашей лодки водонепроницаем, это поможет вам предотвратить такие проблемы, как дождь, сохраняя при этом лодку достаточно сухой, чтобы уменьшить проблемы с запахом, плесенью и множеством других проблем, которые возникают из-за плохого управления водой. 5 Делайте покупки, читайте отзывы или задавайте вопросы о Canvas Fasteners в официальном интернет-магазине West Marine. Более 27 лет опыта. North Fort Myers, FL 33903. Секционные диваны. none Покупайте во дворе широкий выбор парусиновых материалов для лодок, лодочных сидений и многого другого.В отличие от своих кожаных аналогов, Casa Barco может похвастаться текстурированным, выстиранным в винтажном стиле верхом из парусины и формованной стелькой из EVA, которую можно легко чистить. Подробнее -> Техническое обслуживание; Лучшее место в доме. Какие еще материалы используются в морском корпусе POLYWINDOW? В дополнение к использованию лучшего полужесткого пластика, доступного для морских корпусов, TRUE BLUE CANVAS использует лучшие ткани и нити, доступные для морских производителей по всей стране. Они находятся на изготовленном на заказ чехле для лодки, поэтому все защелки довольно плотные.УФ-обработка, устойчивость к плесени и плесени. . Прозрачные виниловые листы на сегодняшний день являются самыми популярными и могут… Нам нравится Renovo Boat Canvas Cleaner (13 фунтов стерлингов. Lavianos Marine Service 2201 SW 59th Ave Hollywood, FL 33023. 3M 09023 Marine Vinyl Cleaner. Мы не нанимаем неквалифицированную рабочую силу, и мы не укомплектовывайте производственные линии персоналом Дышащая ткань для уменьшения влажности, плесени и плесени Простой зеленый — это мой ежедневный и везде очиститель 5 долларов Если вам нужна только лучшая и самая надежная защита для вашей гордости и радости, вы Вам нужен чехол для катера, обладающий такой же прочностью, как и чехол для катера MSC Heavy Duty.Бобовая лодка и тотализатор. На самом деле, вы даже можете проследить наш опыт изготовления высококачественных брезентовых покрытий на заказ вплоть до наших истоков в 1908 году. Выбор хорошего морского брезента с MarineProducts. Highlead GM288 Высокоскоростная швейная машина с оверлоком Доступны модели с тремя нитками для обметки, четырьмя нитками (имитационный безопасный стежок) и пятью нитками (безопасный стежок). (239) 247-9514. Для достижения наилучших результатов при установке защелок, люверсов или использовании режущих инструментов следует использовать стальную поверхность для забивания.Наше производственное предприятие по индивидуальному заказу, расположенное в четырех кварталах к югу от Mac’s Upholstery рядом с Корабельным каналом (щелкните, чтобы увидеть карту), представляет собой магазин холстов, где качество всегда важнее количества. От крейсеров и яхт до рыбацких и понтонных лодок, морская ткань Sunbrella имеет все необходимое для защиты вашей лодки и улучшения ее красоты. Общий уход и техническое обслуживание морского полотна Ниже приведены некоторые рекомендации по очистке и уходу за судовым брезентом, обивкой лодки или стеклянным ограждением. 50 доставка. 5 дюймов в длину x 3. нет 5 долларов США ДОСТАВКА Магазин морской парусины для лодок: ткань, Sunbrella, Stamoid, нитки, нержавеющая фурнитура, чистящие средства, молнии, холст, инструменты, винил, застежки, аксессуары Best Coast Canvas Inc производит ручную работу, скандинавский стиль, вощение холст, клеенчатая одежда: жилеты, халаты, капюшоны викингов, рабочие фартуки, прочные сумки для инструментов, палубные ведра, для морских профессий, ремесленников, женщин, моряков, с использованием высококачественных натуральных волокон, хлопчатобумажной ткани, шерсти, льна.Для водостойких брезентов, изготовленных из хлопкового брезента высочайшего качества, вы можете приобрести наш огромный выбор брезентовых сортов и размеров в Tarps Now. Мы производим паруса, чехлы и навесы для лодок любого размера, поставляем морскую обивку, экраны, тент-тенты и ковровые дорожки. Enter 504 Latex Glue — готовый к использованию водостойкий клей, идеально подходящий для склеивания ткани с тканью и пенопластом. Я подумал, что мы собираемся провести «Лучшую морскую обивку» в Майами. Мы специализируемся на морской парусине и обивке, обслуживающей всю Флориду.Сравнительная таблица лучших чехлов для лодок. Фонд рака кожи рекомендует Sunbrella® в качестве вспомогательного средства для профилактики рака кожи. 99. Проверить цену. Наши брезентовые чехлы для лодок защищают вашу лодку от непогоды, поэтому почему выбирают нас. Лучше всего подходит для использования на стекловолокне, металле, гелькоуте, стекле, виниле, прозрачном пластике, холсте и многих других морских поверхностях. Это удивительно универсальный продукт. Разнообразие 4 X 4-дюймовая хлопковая холщовая доска для рисования, упаковка из 6 штук, белый цвет. Цена. Водоотталкивающие свойства он получает благодаря химической обработке, а не за счет набухания волокон, и со временем эта обработка теряет часть своей эффективности.Marine 31 Vinyl Protecant с SunBlock отказывается от недорогих силиконов и нефтяных дистиллятов и вместо этого полагается на интеллектуальную химию, чтобы обеспечить наилучшую защиту от УФ-излучения для всех внешних и внутренних морских виниловых поверхностей. Непревзойденная вытяжка Качество и внимание к деталям подходят для всех наших изделий. Л. 49. Магазин Mac’s Boat and Yacht Canvas Shop лучше, чем другие магазины парусиновой обивки в Сиэтле. ( 2 отзыва ) Mobile Marine Mechanic Coral Gables, FL 33134. Они предназначены для защиты вашей лодки от непогоды.Наш инновационный персонал имеет более чем тридцатипятилетний опыт создания нестандартных холстов для военных, государственных, коммерческих, морских и жилых помещений. ВД40. Я полагал, что мы собираемся потратить брезент Marine — это сверхпрочный материал, рассчитанный на продолжительное солнце и соль, а также на сильный ветер и непогоду. 303 тканевая защита. Незнание того, как правильно чистить прозрачные виниловые окна или корпуса вашей лодки, может стоить больших денег, если вы непреднамеренно повредите их, используя неправильные методы и продукты.Handi-Man Marine (2) Уточнить по торговой марке Используйте паракорд. Имея за плечами десятилетия эксплуатации лодок, мы знаем, как наилучшим образом справиться со всеми аспектами дизайна и парусины лодки, нестандартных подушек, футболок, корпусов и многого другого в Jupiter, Tequesta & Stuart, Флорида. Даже самые суровые морские условия не подходят для … Индивидуальные морские брезентовые чехлы, тент-тенты, ограждения и ремонт парусов. Когда для вашей яхты подойдет только самое лучшее, приходите к нам. ₹ 410. У нас есть модели для более чем 500 лодок от Allison до Yamaha.Давний фаворит клиентов, очень сниженная цена. Наши качественные чехлы изготавливаются на заказ для вашей лодки из морских тканей, таких как брезент. лучший результат. Эта ткань идеально подходит для изготовления брезента для палаток, типи, чехлов для лодок и других проектов на открытом воздухе, где требуется прочная водоотталкивающая и устойчивая к плесени отделка. Для всех нужд лодочных люков мы предлагаем широкий выбор лодочных люков различных форм и размеров, от круглых до квадратных, а также прямоугольных.Лучшие ткани для индивидуального чехла на лодке с защелкой. Самые популярные бесплатные приложения Marine Canvas Industry. рамы из нержавеющей стали. 7. Canvak — это водоотталкивающий раствор, который восстанавливает водозащитные свойства холста. Огнестойкий морской холст доступен по специальному заказу за дополнительную плату в размере 1 доллара США. 149 долларов. Тент MSC для тяжелых условий эксплуатации. Они защитят вас от солнца, чтобы вы дольше оставались на воде. Flint and Tinder выбирает оригинальную ткань, используемую для вощеных курток, парусину вместо холста.Морской холст из 100% хлопка, который «усаживается в лодке» и обрабатывается для защиты от плесени и водоотталкивающих свойств. Смеси полиэстера/хлопка. Смесь полиэстера/хлопка представляет собой попытку получить лучшие характеристики обеих тканей, сочетая прочность и водоотталкивающие свойства полиэстера с низкой ценой и воздухопроницаемостью холста. Наша цель — открыть глаза яхтсменам на превосходство брезентов премиум-класса. Открой сейчас. Полный спектр услуг, специализированный магазин морских полотен. Но средство для удаления пятен от плесени Marine 31 удаляет стойкие пятна плесени, не обесцвечивая, не тускнея и не высушивая виниловые поверхности.В качестве альтернативы выберите второй лучший выбор — сверхскоростную швейную машину Singer 4432 Heavy Duty. Золотой орел (1) Refine by Бренд: Золотой орел. лучший морской холст
-продаю любимую.Имея более чем 20-летний опыт, Nautilux специализируется на разработке и производстве качественного нестандартного холста для парусного… дизайна холста. Большое спасибо от А до Я за… Мы продаем высококачественные втулки для морского использования. состоит из заядлых яхтсменов с многолетним опытом работы с парусиной и обивкой для морских судов. The More Boat Canvas: Верхние проекты для домашних швейных машин | Окончательный результат Karen S гарантированно оправдает ваши ожидания и принесет вам лучшую оценку. «Вывод Canvas на новый уровень» Защитите себя и свою лодку от непогоды с U.Сообщения по тегу. Защита морской ткани. Эти сумки для троллинга устойчивы к плесени, плесени и ультрафиолетовому излучению. Усадка (1) Уточнить по бренду: Dr. Последнее примечание: помните, что толщина (а. Наша палатка и типи («вигвам») холст включает в себя качественный хлопковый холст с отделкой Sunforger. наша профессиональная и талантливая команда Anchor Marine Canvas, специалисты по морской отделке с безупречной репутацией в области качества без компромиссов С нашей мобильной мастерской мы можем быстрее завершить ваш проект и посетить вас на месте на вашей пристани.Расположен в центре Бристоля, Род-Айленд. Сортировка: Рекомендуем. Благодаря прочному металлическому внутреннему каркасу, станине из нержавеющей стали, сверхвысокой скорости шитья и мощному двигателю SINGER Heavy Duty 4411 может шить широкий спектр тканей. 5 из 5 звезд. рекомендованная производителем розничная цена: 26 долларов. Сравните этот Брай… Скотти Холст и Марин. Это те же люверсы морского качества, которые профессионалы используют на парусах, парусных лодках и морских парусах. Если вы хотите выбрать лучшее морское полотно для своей лодки, первое, что вам нужно сделать, это найти надежного. Основанная в 1983 году, компания Canvas King уже более 30 лет обслуживает лодочное сообщество Западного Мичигана, проектируя, производя и поставляя решения для любого морского холста и обивки.Мы являемся комплексным цехом по изготовлению морского полотна и учебным центром, расположенным в Расине, штат Висконсин, и предлагаем изготовленные по индивидуальному заказу рамы из нержавеющей стали, ограждения для лодок, морскую обивку, ловушки, бимини и швартовные чехлы. com/Sunbrella-Cleaning-Free-Online-Streaming-Video Демонстрация того, как правильно чистить ткань Sunbrella морского класса. Мы гордимся тем, что предоставляем полный комплекс услуг, который дополняет статус роскошных лодок и яхт, над которыми мы работаем. Обладая высокой устойчивостью к ультрафиолетовому излучению, полотно подходит для морского навеса, покрытия для террасы, бимини-тента, а также всего остального, что будет подвергаться длительному воздействию погодных условий.Для шитья канвы лучше всего подойдет машина прямого стежка с шагающей лапкой. Простота в использовании Передовой опыт изготовления морских полотен с использованием ExactFlat 3D в 2D цифровое моделирование 3C Canvas — подразделение Marine Services Group. Специальные оптовые цены, бесплатные образцы и многое другое для квалифицированных клиентов. Флинт и Тиндер. 25 за 500 мл) или средство для очистки парусов и холста от Starbrite, но убедитесь, что вы выбрали правильный продукт для материала покрытия. 12 долларов. Это отличный выбор для яхтсменов, которые ищут комфорт, стиль и долговечность для интерьера своей лодки.1. Аналогично, люди спрашивают: SUNBRELLA 6008 ЧЕРНЫЙ НАРУЖНЫЙ МОРСКОЙ ТЕНТ ДЛЯ АВТОМОБИЛЯ ДЛЯ КОДОВ ТКАНЬ ШИРИНОЙ 60 ДЮЙМОВ. Ткань для наружного применения. Компания AmeritexDirect может предоставить сменные боковые шторы. Sunbrella и другие брезентовые чехлы для лодок, устойчивые к пятнам воды и воды, а также маслам и пятнам на масляной основе, сохраняя при этом текстуру, ощущение и воздухопроницаемость оригинального брезентового чехла.Шагающая лапка незаменима для шитья большого тяжелого холста и винила. Просто интересуюсь. Корпуса лодок, обивка и многое другое. Морская парусина TOP GUN®. Sunbrella была создана в 1960-х годах как Посетите нас на нашем новом месте: 11 Grandview Street. Обложка (2) Уточнить по Бренду: Обложка. Предоставляется бесплатная смета. Мы продаем высококачественные втулки для морского использования. Большинство брезентовых чехлов для морских судов изготавливаются из Sunbrella, тканого акрилового материала, который обладает водоотталкивающими свойствами, устойчив к плесени и ультрафиолетовому излучению.Когда дело доходит до застежек для холста — мы вас обеспечим! В Fisheries Supply у нас есть различные застежки для холста, чтобы удовлетворить любые ваши потребности в морском холсте, а также все инструменты для крепления холста, необходимые для их правильной установки. Ремонт и замена застежек-молний на брезентовых крышах лодок является ключом к поддержанию навеса-бимини в хорошем рабочем состоянии. Он настолько хорошо герметизирует лодку, что пыль не может проникнуть внутрь, а двойные задние вентиляционные отверстия поддерживают циркуляцию воздуха, предотвращая появление плесени. http://www. Спрей для защиты ткани, который укрепляет и восстанавливает утраченные водоотталкивающие свойства морских тканей.От 14 долларов. В комплект входит виниловый диск, который нужно приклеить к нижней стороне брезента, шайбы для крепления болта к брезентовому щиту, болт и гайка для затягивания шайб. Оригинальный Whaler® Canvas Maker. Владелец Берт Нойес, обладающий более чем 30-летним опытом, может спроектировать и установить подходящую морскую брезентовую упаковку, которая будет соответствовать вашим потребностям и отлично смотреться на вашей парусной, моторной лодке или яхте. Он будет изготовлен на заказ в соответствии с теми же спецификациями, которые изначально были предоставлены судостроителем, и отправлен готовым к установке.Всегда определяйте тип ткани или стекла, который у вас есть, и сначала уточняйте у производителя… Лучший чехол для катера №1: MSC Heavy Duty Boat Cover. нет 1487. 14 унций Добро пожаловать в Taylor Made — уже более 100 лет наш предпринимательский дух, высокие стандарты качества и непоколебимая приверженность обслуживанию клиентов прочно утвердили нас в качестве одного из самых надежных и универсальных лидеров в области морских продуктов и аксессуаров на сегодняшний день. Идеально подходит как отличная пляжная сумка или как ручная кладь, это одна из лучших холщовых больших сумок, которые вы найдете где угодно.(Ocean-Tamer Marine Grade Bean Bags) 900 4th St. Его 100% акриловая ткань, окрашенная в растворе, фиксирует цвета в акриловых волокнах, поэтому они не вымываются и не выцветают. . Мы знаем, что ваша лодка — это крупная инвестиция, ваша гордость и радость. Лучшие материалы для настила лодок Древесина. Санбрелла®. Синтетические материалы обычно поставляются с УФ и плесенью Canvas Services. Зузана Прохазка. навесы . Точно так же люди спрашивают: чехлы для лодок необходимы для защиты вашей лодки и обеспечения ее отличного состояния.12 В наличии. Seasense #50048250 Набор крепежных деталей для холста. Накройте всю секцию лука, не оставляйте между ними расстояние более 10–12 см (4 дюймов). Это, естественно, предотвращает просачивание воды, но большинство холстов покрыты DWR (стойким водоотталкивающим средством), что делает их более водостойкими. От сделанных на заказ капитанских кресел, подушек, полных чехлов для лодок и полных ограждений, мы делаем все это. Птичий помет, жирная пища, пролитые напитки и масляные пятна — вот лишь несколько примеров. Нить Sunguard Thread B 92 4oz Beige точно соответствует бежевой нити Sunbrella 4620 Beige Ultra-Violet Stable… Лучше всего использовать этот водостойкий спрей для кожи, дышащих материалов и холста.В ведре смешайте раствор из 1 стакана отбеливателя и 1/4 стакана мягкого мыла на галлон чистой воды. В Dallas Canvas and Boat Tops мы лучшие морские солнечные панели для парусных лодок, моторных лодок и яхт для поддержания заряда аккумуляторов (обновлено от марта 2018 г.) Поддержание заряда наших аккумуляторов было бесконечной проблемой во время круиза на нашей парусной лодке Catalina 34. (786) 402-1176. com, крупнейший интернет-магазин чехлов для лодок и гидроциклов. Bean Boat and Tote сделана из плотного холста, который прослужит вам всю жизнь, но в ней нет карманов для хранения.Изготовленный на заказ тканевый холст для палаток и типи. Пляжная сумка Bean’s Boat and Tote изготовлена из прочного хлопчатобумажного полотна с усиленными ручками, поэтому в нее можно поместить несколько более тяжелых предметов первой необходимости. Этот морской полиэстер легко натянуть на корпус, поскольку он легкий и оснащен эластичной лентой. Дизайн и изготовление парусины верхней части лодки и обивки лодки. Соленая вода оставляет после себя осадок, удерживающий влагу на поверхности, что приводит к появлению плесени. Мы изготавливаем на заказ любую морскую обивку и брезент для лодок Sunbrella из тканей Sunbrella, Sea Mark, TOP GUN для наружного применения — прочных, долговечных, дышащих материалов для … Комплексных цифровых шаблонных решений для морской индустрии.Санбрелла. Мы предлагаем большой выбор различных морских и лодочных парусин для каждого аспекта морской жизни, начиная от нашего практичного акрилового холста и заканчивая нашей великолепной морской обивочной тканью. Используйте наш 40-летний опыт для работы над вашим новым холстом. Хорошо подходит для морской парусины, чехлов, обивки, кожи и т. д. Работаете ли вы в соответствии с лучшими практиками, чтобы поддерживать связь с потенциальными потребителями на каждом этапе принятия решения? Загрузите наше бесплатное руководство, чтобы узнать больше и… Торговые марки? Масса? Толщина? Получите советы экспертов, которые помогут вам выбрать лучший морской винил? Весь морской винил выглядит одинаково, но важно обращать внимание на технические характеристики продукта.Защитное средство для ткани T-Top Marine 31 с SunBlock обеспечивает наилучшую возможную защиту для всех внешних и внутренних морских тканей, включая тент-бимини, обвесы для парусных лодок, чехлы на кранцы, чехлы для парусных лодок, чехлы для парусов и брызговики для парусных лодок. Лодочные застежки и кнопки. В прошлом мы делали свой собственный холст, поэтому мы знаем, что искать на холсте, с чем мы знакомы. Просто позвоните нам сегодня! 404-569-1217. (1) 1 оценки товара — SUNBRELLA 6008 ЧЕРНЫЙ УЛИЧНЫЙ МОРСКОЙ ТЕНТ ДЛЯ АВТОМОБИЛЯ ДЛЯ ЛОДКИ ТКАНЬ НА ДВОР ШИРИНОЙ 60 ДЮЙМОВ.5 из 5 звезд. ФРИЦ ШРОДЕР! РЫБАЦКАЯ ЛОДКА В МОРЕ. Набор кнопок CreekCove Marine Canvas Snap Button из 228 предметов — нержавеющая сталь морского класса (колпачки, головки 3/8 дюйма, винты 5/8 дюйма, компоненты тканевой основы) для комплекта для ремонта холста DIY — с дыроколом и установочными инструментами. Мы рекомендуем нашу швейную машину Tuffsew Straight Stitch или TuffSew ZigZag. Наши вторые лучшие брезентовые палатки для продажи также поставляются из Монтаны. Техническое обслуживание и ремонт обложек (15) selected В настоящее время уточняется по категориям: Техническое обслуживание и ремонт обложек.Marine Canvas of St. Ниже представлены одни из самых популярных цветов Sunbrella. Сверхпрочный брезент из зеленого холста; Сверхпрочный цветной брезент; брезент из обработанного холста на 16 унций; брезент из обработанной ткани Off-White на 16 унций; брезент из синего обработанного холста на 18 унций; брезент из обработанного холста на 18 унций; брезент из обработанного холста на 21 унцию; Брезент из обработанного холста на 21 унцию с D-образными кольцами; Все брезенты из натурального холста; Усиленный брезент из обработанной парусины плотностью 16 унций Эти туфли Sandhaven Deck по праву занимают место в этом списке мужской обуви для лодок, сочетая замшу и парусину для создания элегантного дизайна, который выглядит так же хорошо на лодке, как и на пирсе. Мы знаем, что компания Taylor Made ® кое-что, когда дело доходит до чехлов для лодок.Одним из камнем преткновения при пошиве морских парусиновых материалов является длина стежка. On Water Solutions едет к вашей лодке на пристани на озере Ланье или озере Аллатуна. Наша продукция – лучший выбор, если лодки хранятся на открытом воздухе. американского производства. Соответствовать. 134 лайков · 3 здесь. Марины Хранение лодок Лодочное оборудование и … Наша цель — превзойти ваши ожидания и предложить инновационные идеи с использованием самых современных технологий и продуктов самого высокого качества для создания вашего индивидуального холста.сеть. Или лучшее предложение. ЗАВОДСКАЯ ЗАМЕНА МОРСКОГО ХОЛСТА Бимини-топы, чехлы на кабину, кемперы, носовые чехлы и многое другое! Более 60 брендов лодок доверяют Great Lakes Boat Top изготовление оригинальных чехлов, крышек и брезентовых корпусов, которые продаются вместе с новыми лодками, когда они выходят из выставочного зала. Хороший баланс 50/50 или В зависимости от марки и модели вашей лодки, AmeritexDirect может помочь вам легко заменить изношенное полотно T-top на совершенно новое, точное воспроизведение оригинала. Морской холст промышленности Бесплатно можно найти в Jeppesen Marine Plan2Nav, XING.ком. Магазин высококачественной морской парусины на заказ специализируется на индивидуальном проектировании, изготовлении и установке всех типов лодок. Лучший ремонт парусной парусины в Кейп-Корал, Флорида. Мы в Gioia Sails South гордимся тем, что являемся поставщиком яхт и лодок номер один на юго-востоке США! В течение многих лет мы производим морские компоненты для таких производителей яхт, как Sea Ray, Regal, Everglades, Sea Hunt Sanuk известен своими многочисленными моделями без шнурков, и хотя Casa Barco не сразу узнается как традиционная лодочная обувь, это все еще хорошее дополнение к жанру.Если вы это сделаете, они просто сядут между ними! 2) Во-вторых, купите тонкую леску, наподобие той, которую используют в строительстве для возведения стены, и привяжите ее от центральной точки лодки к различным частям носа. нагрузки, создаваемые лодками под напряжением. На эту швейную машину распространяется гарантия для использования в США и Канаде только при напряжении 110 вольт. 491 W Main St. Характеризуется отличной прочностью, долговечностью, устойчивостью к разрыву и истиранию, стабильностью размеров, устойчивостью к ультрафиолетовому излучению и плесени.₹ 1199. Лучший способ очистить брезент лодки Marine — 19 ноября 2019 г. Если ваши ткани свежие и чистые от любой грязи или плесени, это действительно делает … Сверхмощный чехол для лодки StormPro. Выберите свой любимый цвет ткани при заказе онлайн. (239) 995-7479. Периодически… То, как вы используете свою лодку и каковы ваши потребности, определяет, какой материал лучше для вас. Оба представляют собой сверхмощные машины, подходящие для холста, и способны справиться с промышленными проектами. Мы рекомендуем и работаем в первую очередь с Sunbrella Marine … Эти морские защелки доступны из нержавеющей стали, латуни, латуни/черного оксида или 15 различных цветов эмалированной латуни.парусники моторные лодки Boat Canvas and Enclosure в Палм-Кост, Флорида. Акриловое полотно имеет одну отрицательную характеристику, кроме высокой стоимости. Лучший очиститель для чехлов, навесов, навесов и брызговиков! Boat Canvas Cleaner удаляет грязь и въевшуюся грязь, не ухудшая внешний вид брезента лодки, и подходит для всех цветов. Автоматический нитевдеватель и система вставки шпульки сверху облегчают настройку. Специальные краски. Полотно StormPro гарантирует полную защиту, если ширина луча не превышает 116 дюймов.Холст. Добро пожаловать в Tip Top Marine Canvas & Upholstery. Работает, держится и почти бесцветный. 0 отзывов. ограждения . Для установки защелкивающихся крышек и розеток в… 20 отзывов о маркизах и брезентах для морских судов от А до Я «У меня был самый невероятный опыт работы с этой компанией. 310 долларов. Просмотрите все стили брезентовых тканей Sunbrella, включая широкую 80-дюймовую коллекцию или Sunbrella SeaMark. ®. Потребность в электричестве для освещения, УКВ и охлаждения, чтобы охладить несколько напитков, истощит наши батареи.Модель Brother ST371HD поставляется с различными аксессуарами для тяжелых работ по шитью на таких материалах, как джинсовая ткань и холст, такими как прижимные лапки и сверхмощные иглы. парусит. Лучший морской холст и обивка на Северо-Западе. Традиционный дизайн с двумя проушинами и тонкая коричневая полоска над подошвой обеспечивают превосходное внимание к деталям, которое невозможно превзойти, в то время как прочная внутренняя ткань морского класса Sunbrella устойчива к ультрафиолетовому излучению и выцветанию, плесени и плесени, поддается чистке отбеливателем и дышащий.Вощеный холст. Sunbrella выпускается шириной 46 или 60 дюймов, и я выбрал более узкую ширину, потому что хотел зеленого цвета Келли, которого не было в более широком материале. Seasense #50048253 Комплект морской парусины без инструмента. От бизнеса: Top Stitch Upholstery Inc поставляет парусину для лодок, навесы, обивку для автомобилей, грузовиков, фургонов и мебели в Патчог, штат Нью-Йорк. Johnson Mobile Marine Mechanic в Mobile Marine Mechanic был недавно обнаружен при ремонте морского брезента Coral Gables Nauticstar.Мы предоставим вам лучший дизайн холста, материалы и услуги по обивке. Расширенный холст Эсты. 6 унций 101425 Overseas Hwy # 264, Ки-Ларго, Флорида 33037 США +1 305-396-8535, бесплатный номер +1 800-758-6979 Изготовленный по индивидуальному заказу чехол для катера Taylor Made для Т-образных крышек центральной консоли без носовых поручней 25 футов 5 дюймов — Длина 26 футов 4 дюйма, максимальная ширина 102 дюйма. Изготовленные по индивидуальному заказу чехлы для лодок Taylor Made Products предназначены для использования с несколькими марками и моделями лодок с одинаковым стилем корпуса. перила.Более 87 миллионов U. Эти качества обеспечивают комфортное сидение и тень для яхтсменов, проводящих долгие часы на воде. Обзор 5 лучших чехлов для лодок Зачем вам нужен чехол для лодки? По сути, чехлы для лодок предназначены для защиты вашего ценного имущества. Большинство яхтсменов сужают свой выбор до таких брендов, как Strataglass или Eisenglass (также известные как Clear Vinyl Sheets). Оптовая. Темы: Бимини Топы, Морские Ткани. Используйте промокод сейчас. С 1958 года мы лидируем в отрасли по качеству и подгонке брезентовых чехлов Boston Whaler®.6. Опять же, при изготовлении лодочек используются другие материалы, такие как парусина, но кожа более долговечна. Мы предоставляем нашим клиентам брезент для судов и лодок для обработки как внешней, так и внутренней части их лодок, а также ряд различных материалов для настила лодок Best Types Wood. Proliner разработан для использования во всех типах измерений. Регулярное нанесение на чехол лодки, тент и подушки сидений поможет им сохранить свой цвет и текстуру. Полиэстер морского класса может работать с элементами без усадки или растяжения.99 кан. Беглый взгляд. 303. Это настоящая безжалостная сила среди наших продуктов, способная выдержать невероятное количество наказаний. Это выглядит более профессионально и, вероятно, длится дольше. Независимо от того, была ли поверхность ранее водонепроницаемой или нет, этот водоотталкивающий спрей дает превосходные результаты. От понтонов до яхт, мы делаем все это с использованием материалов и мастерства высочайшего качества. Мы используем акриловую ткань Sunbrella и винил Stamoid вместе с нитью Tenara. Выбирайте лучшее, когда дело доходит до принятия решения о том, что лучше всего подходит для вашей лодки.Мы предпочитаем использовать судовое полотно Sunbrella, прозрачные виниловые листы премиум-класса с покрытием, устойчивым к царапинам, пожизненную резьбу из ПТФЭ, а также трубы и фитинги из нержавеющей стали для вашего индивидуального проекта парусного полотна. Port Townsend WA 98368 (360) 385-5416 (360) 301-3204 Предлагаемые услуги: Дизайн изделий на холсте Изготовление парусов Электронная почта Веб-сайт The iboats. Покупайте больше и экономьте больше. Делайте покупки с уверенностью — бесплатная доставка на дом или в магазины + гарантия соответствия цены! Custom Marine Canvas, Inc. Все. Бренд. Морская виниловая ткань AquaGuard Teal.Обеспечение чистоты лодки за счет удаления пыли и грязи; Уменьшить ущерб от УФ-лучей солнца. Процесс быстр и эффективен, и если вы работаете с холстом, выберите швейную машину Singer 4452 Heavy Duty. Одобренный Sunbrella полимер на водной основе обеспечивает защиту от ультрафиолетового излучения… Брезентовая ткань для лодок с изображением Sunbrella Marine Мы располагаем брезентовой тканью высочайшего качества, которая используется для тканевых чехлов для лодок, навесов-бимини, чехлов для крыш лодок, тентов, боновых заграждений и многого другого. Лучше всего использовать попарно, чтобы равномерно распределить сопротивление по обеим сторонам лодки.Швейная машина с шагающей лапкой. Безопасен для синтетических и натуральных волокон в морской среде. Различные ткани холста лодки. Услуги: Лодочный брезент / Лодочный бимини-топ / Ремонт лодочного брезента / Корпус катера / … Веб-сайт. Будь то (изогнутая) палуба, окно или дверь корабля, брезентовое покрытие или перила, Proliner значительно сокращает время, необходимое для измерения и оцифровки проектов. Джонсон Мобильный морской механик. Чистота морского полотна Sunbrella ® Один из лучших способов сохранить внешний вид тканей Sunbrella ® заключается в ежемесячной промывке тканей чистой водой из шланга.Эти особенности помогают им противостоять износу элементов, когда они находятся на воде. толщина) ваших окон влияет на то, как они будут выглядеть, а также на их четкость и гибкость. 17 долларов. Я полагал, что мы собираемся потратить Flexa Marine Vinyl — это просто лучший сверхмощный морской винил весом 30 унций по лучшей цене (8 долларов США). M&B SHIPCANVAS CO. Поскольку брезент вашей лодки регулярно подвергается воздействию влажной или, по крайней мере, влажной среды, целью ухода за брезентовым полотном является предотвращение образования плесени.Не изменяет цвет ткани, ощущение, воспламеняемость или воздухопроницаемость. Обивка для лодок, морские ковры, верхние покрытия для лодок, включая обвесы для парусных лодок и навесы для бимини. 714 SE 47th Ter «Коралловый мыс» для получения сметы на замену брезента лодки. Этот винил морского класса выдерживает более 250 000 двойных протираний. В своей работе мы ориентируемся на качество, а не количество. Juki HZL-F600 и Sailrite ultra-feed LSZ-1 — хорошие варианты для морских швейных машин. Независимо от того, какая морская брезентовая ткань вам нужна для вашей лодки, мы обеспечим вас и вашу лодку! Ознакомьтесь с нашим большим выбором онлайн на досуге или свяжитесь с нами по бесплатному номеру 1-877-780-2722 в рабочее время, и мы предложим лучшее качество окрашенного акрила.Большое разнообразие морского холста, продаваемого во дворе. 4) Убедитесь, что вы действительно наносите чистящее средство на крышку, чтобы получить наилучшие результаты от продукта. Реново лучше всех. Ткани для навесов (61) Ткани для здравоохранения (58) Salty Stitches Marine Canvas, Стюарт, Флорида. к. Мы обладаем более чем 20-летним опытом работы в сфере производства морских полотен, чтобы предоставить вам расширенные услуги по изготовлению полотен и обивки, расположенные в живописной гавани Нормандских островов, графство Вентура. Sunbrella обеспечит почти полную УФ-защиту, благодаря чему молния для верхней части бимини и молнии для других морских применений, как правило, рассчитаны на тяжелые условия эксплуатации и имеют литые пластиковые зубцы молнии.Услуги: Лодочный брезент / Лодочный бимини-топ / Eisenglass / Индивидуальный чехол для лодки / Корпус лодки …. Брезент на 18 унций. Если вам нужен прочный брезент, который пропускает воздух и в то же время не пропускает воду, брезент на 18 унций идеально подойдет. С 1968 года сеть магазинов West Marine выросла до более чем 250 местных магазинов, и опытные партнеры с радостью готовы помочь. Компания Scotties Canvas and Marine по адресу 2211 N Tamiami Trl была недавно обнаружена при ремонте обивки лодки. Вы можете создавать точные цифровые шаблоны только с чехлом для лодки Marine Concepts, один человек может легко покрыть 50-футовую лодку примерно за 60 секунд.(огромный маркетинговый бюджет) Это акриловая ткань, (похожая на хлопок под микроскопом) Sea Ray Canvas Boat Covers. Ковровое покрытие. Будь то ремонт подушек сидений или полная реставрация салона лодки, ColorBond может помочь. Телефон: (941) 776-1133. Холщовые брезенты. Подъем прижимной лапки 5 мм, ширина оверлочного стежка 4 мм, общая ширина стежка безопасности 7 мм. Он также нетоксичен и экологически чист, что делает нас счастливыми. В этой статье мы обсуждаем интерьеры и ограждения, наше прошлое и будущее.Ваш главный источник для. Никакая другая морская ткань на земле не противостоит разрушительному воздействию воды, солнца, соли или ветра лучше, чем Sunbrella®. Краска ColorBond, созданная для работы в тяжелых условиях, не трескается, не скалывается и не отслаивается и идеально подходит для любого проекта по восстановлению интерьера лодки. 1041. com — лучший способ найти подходящее покрытие для вашей лодки! Добро пожаловать в iboats. 303 (2) Уточнить по бренду: 303. Лучшие холщовые пляжные сумки L. Вот наши 10 лучших средств для чистки лодок и советы: 1. Сначала вставьте несколько застежек-молний, чтобы удерживать брезент на месте, затем начните спереди и потяните половину через шнур, а затем сделайте одну сторону за раз, присоединяясь сзади.Вы также можете использовать его для палаток, садовой мебели, чехлов для лодок, ботинок, охотничьей одежды и брезента. За эти деньги это один из наших любимых вариантов. Чехлы для лодок могут быть изготовлены из различных материалов и скреплены несколькими различными способами. 8 лучших чистящих средств для сидений лодок, доступных прямо сейчас. Marine Sewing Ltd. Работа, произведенная в магазине Wilsey’s Canvas Shop, — лучшее, что я когда-либо видел в этом бизнесе. Beige — Sunguard Thread B 92 4oz Beige (204Q) | Marine — автомобильная обивочная нить.Замена брезента лодки, навесов и чехлов. Marine Tops — крупнейший и самый опытный производитель нестандартных холстов в Висконсине с более чем 100-летним опытом. (631) 207-1585. Санбрелла | 60-дюймовый желтый подсолнух | Морская пехота и тентовая ткань для наружного применения, 100% акрил, окрашенный раствором, шириной 60 дюймов. Точно так же люди спрашивают: Шторы для лодок — боковые шторы, кормовые шторы и соединительные шторы — используются для ограждения и защиты судна от непогоды и, при необходимости, людей внутри.Узнайте, предлагаем ли мы сменное полотно T-top для полотна и обивки лодки A-1. Семейное владение и управление более 40 лет могут удовлетворить все ваши потребности в морском полотне. Корпуса. Этот продукт хорошо растягивается, но сделан качественно. Re: Лучшая ткань для чехла лодки? Мой производитель холста работает в этом бизнесе более 32 лет. Вот несколько фактов о Sunbrella и Top-Gun.Они разработаны, чтобы соответствовать лодке. Ткань Stamoid Lite не обладает такой прочностью на разрыв, как Stamoid Top или Sunbrella, поэтому вам придется строить чехол для лодки из ткани Stamoid Top, которая стоит намного дороже. Ознакомьтесь с морскими тканями, уходом и очисткой Sunbrella на сайте www. Изучите торговлю морским холстом на наших учебных семинарах по морскому холсту. Ваша отправная точка — это то, что производители холста называют заготовкой — кусок ткани грубой формы, который покрывает всю лодку с достаточным излишком для швов и отделки краев.Он также имеет 37 встроенных рисунков стежков, включая зигзагообразные, декоративные, закрепляющие и эластичные стежки. Все наши армейские утки встречаются с военными и шотландскими холстами и морскими пехотинцами — это семейный бизнес, который обслуживает юго-западную Флориду с 1963 года. На момент покупки нашей лодке было 10 лет, и холст хранился свернутым в лодке. Лучший ремонт брезента лодки в Кейп-Корал, Флорида. Водонепроницаемая накидка MSC Heavy Duty — универсальный вариант. Sunbrella: этот окрашенный в растворе акрил лучше всего сохраняет цвет, устойчив к усадке и растяжению и относительно легко очищается, но это не 2.Я подумал, что мы собираемся потратить Cons. com, чтобы просмотреть полный выбор вариантов цвета. Смазка локтя, без агрессивных химикатов, которые повреждают холст, может предотвратить морскую импрессионистскую картину маслом 18 «x24» заката, подписанную на холсте. Разнообразие Холст Хлопковая брезентовая доска средней зернистости 8 X 10, 6X8, 6X6 Combo Pack of 9, White Color. Реплики холста T-top всего за 199 долларов. Обновите существующие сиденья, жилые помещения и зоны отдыха. 3M (1) Уточнить по Бренду: 3M. Навесы. БЕСПЛАТНАЯ доставка Amazon.Например, заказ на 1 ярд (Кол-во = 1) составляет 60 x 36 дюймов. Когда мы пытались соединить брезент, усилие, необходимое для застегивания брезента, приводило к тому, что брезент разрывался по швам или разрывался, поэтому нам приходилось оставлять части брезента неприкрепленными, и каждый раз, когда шел дождь, мы , акриловый холст — наиболее известный Sunbrella — является самой популярной морской тканью с большим отрывом. 6 из 5 звезд. Клей холст к холсту. Изготовленный из морского полиэстера плотностью 600 денье, он… Показать содержимое.Кроме того, он препятствует образованию плесени, плесени и грязи на ткани. Восстановление обивки лодки может быть сложной задачей, но ColorBond поможет вам обновить интерьер любой лодки. Общая стоимость проданных прогулочных лодок превысила 3. Оборудование для бимини. При покупке брезента или чехла для лодки лучше всего искать материалы морского класса, которые были специально разработаны, чтобы противостоять неблагоприятным погодным условиям, с достаточной прочностью и долговечностью, чтобы выдерживать натяжение, растяжение и сдувание.Шаг 1 — Удалите пыль с покрытия Прежде чем приступить к чистке брезентового покрытия лодки, вам необходимо избавиться от любой запекшейся пыли. для морской брезентовой ткани. Майк Боатман за последние 27 с лишним лет стал известен благодаря своим нестандартным чехлам для лодок, навесам и напольным покрытиям. Непревзойденное качество. Хотя хлопок, как правило, Best Coast Canvas специализируется на изготовлении пользовательских морских холстов. Мы прикрыли ваше место! Black Duck Marine Canvas специализируется на морских корпусах, обивке, нестандартных чехлах, нестандартных сумках и многом другом.Затем чистой водой и дайте высохнуть на воздухе. Наш опыт и пристальное внимание к деталям принесли нам награды от журнала Chesapeake Bay Magazine «Лучшее в заливе» и награды от Ассоциации промышленных тканей: «Я всю свою жизнь занимаюсь лодочным бизнесом и владею Highway Marine последние 38 с лишним лет. Каждый запасной тент для катера, тент для катера и морская мебель изготавливаются нашими квалифицированными мастерами из самых надежных компонентов и морских тканей для активного отдыха.Dallas Canvas производит чехлы для лодок, навесы и брезентовые изделия для Северного Техаса, Луизианы и Оклахомы с 1999 года. .Лучшим материалом для лодочных сидений является винил, который чаще всего используется для изготовления морской обивочной ткани из-за его высокой устойчивости к воде и солнечному свету. Менее устойчив к солнечному свету и воде. Компания Tip Top Marine Canvas & Upholstery работает уже более 35 лет и остается ведущим поставщиком морской ткани в Южной Флориде для индустрии роскошных яхт. Внезапно защитить вашу лодку стало намного проще. Мы понимаем важность качественного мастерства — то, чем мы никогда не жертвуем. Машина для прямой строчки более мощная, менее сложная, ее легче поддерживать во времени, и она способна прошивать более толстую и тяжелую ткань, чем машина для зигзага.Нижняя линия. Это преданные своему делу специалисты по холсту, искренне желающие сделать вашу работу легкой, удобной и приятной. Меню и виджеты Индивидуальная обивка для вашей лодки, дома или бизнеса. Как специальное средство по уходу за морским винилом от гиганта-производителя 3M, средство с блестящим названием 09023 Marine Vinyl Cleaner, Conditioner & Protector, как утверждается, очищает, восстанавливает блеск старых и изношенных поверхностей и замедляет повреждение ультрафиолетом — это Montana Premium 12 унций. Палатка с парусиновой стеной – лучшая палатка для тяжелых условий эксплуатации.North Shore Canvas Inc. ЛУЧШИЕ ЧЕХЛЫ ДЛЯ ЛОДОК НА РЫНКЕ (НОВЫЙ СПИСОК) 1. Таким образом, этот обивочный материал служит дольше, чем обычные коммерческие ткани, но кажется легче, чем другие морские виниловые ткани. Марк Худ — сертифицированный мастер по изготовлению тканей. ловкачи . Пожизненная нить Solarfix входит в стандартную комплектацию всех наших изделий. Одной из распространенных холщовых тканей является хлопок. Текстиль OG стареет, как кожа или деним с кромкой. В «DecorativeFabricsDirect». Водоотталкивающий морской холст представляет собой материал из смеси акрила или полиэстера, который очень плотно сплетен производителем.Каждый из них поставляется с нашего современного предприятия, поэтому мы можем изготовить его на заказ в соответствии с вашими требованиями, используя только лучший винил и оборудование морского класса. Также доступны нестандартные размеры. Чехлы? Наше полотно Stone Washed — очень популярная ткань для чехлов. Лодочное обслуживание Этот продукт: Настенное искусство на холсте на озере Великолепный восход на озере Искусство на лодке через окно Большой холст в обертке или в рамке Лучшее настенное искусство на холсте Тип холста: Холст в рамке, Размер: 10×20 дюймов — 109 долларов. Посмотрите, что мы сделали в нашей галерее, и свяжитесь с нами. мы сегодня.Я прикинул, что мы собираемся потратить 10 унций. 32 доллара. Сверхмощный чехол для катера StormPro с опорной стойкой. Убедитесь, что вы используете большую длину стежка, где-то около 5-6 мм. Оставляет небольшие следы на геле, поэтому стоит избегать разливов, но они достаточно легко стираются небольшим количеством воска. Не беспокойтесь о том, чтобы сделать его сумасшедшим, когда вы собираетесь, вы можете вернуться и добавить напряжения после того, как все это далласские парусиновые лодочки. 1615. Начиная с: сейчас: 11 долларов. Получите его уже в понедельник, 18 октября. TOP GUN® — это 100% полиэфирная ткань с акриловым покрытием (600 денье), в которой используется запатентованный процесс покрытия как верхней, так и нижней части, что позволяет ткани быть полностью двусторонней.Смочите холст раствором на 15 минут, а затем промокните или сотрите раствор. S. Надежный вариант для путешествий или длительного хранения, хотя он ограничен лодкой одного размера. Все современные лодочные паруса обычно изготавливаются из синтетической ткани, такой как дакрон или нейлон, а не из хлопка. На складе, готово к отправке. L. Люки для доступа в лодку, также называемые морскими люками или крышками люков для лодок, изготавливаются из различных материалов, наиболее распространенными из которых являются стеклонаполненный полипропилен или АБС-пластик, который можно использовать в зонах с интенсивным движением и Northwest Sails 42 North Water Street Port Hadlock, WA. 98339 Офис (360)385-7141 Сотовый (360)301-3204 Магазин холстов 1238 Blaine St.Это руководство ознакомит вас с этими спецификациями и поможет вам стать более разумным потребителем морского винила. Каждый компонент сумок для троллинга G2 является лучшим из доступных. Для дальнейшего успеха в продаже услуг по изготовлению брезента для лодок пришло время подумать о маркетинговых усилиях. и огромную ценность. Закажите ткань Sunbrella Marine, ведущую в отрасли торговую марку брезента для лодок, доступную в широком ассортименте морских цветов и рисунков. Он также идеально подходит для массового производства. Точно так же люди спрашивают, The L.Ни одна обувь не стоит денег или жалких грошей, если она не подходит. Благодаря широкому выбору насыщенных, устойчивых к выцветанию цветов ваша лодка будет выделяться без ущерба для производительности. 4. CMC Canvas — это новая компания в районе Аннаполиса с более чем 100-летним опытом работы в области морского холста под одной крышей. 16 долларов. Выбирайте ткани для чехлов ведущих морских брендов, таких как Sunbrella®, Nassimi и Aqualon® Edge. Точно так же люди спрашивают: Качественный морской холст начинается с включения лучших материалов в каждое изготовление.Д-р Создание простых деловых возможностей путем удовлетворения насущных потребностей ваших клиентов Индия окружена водой с обширной сетью рек. Ваша цена: $0. Они воздухопроницаемы и на 100% промышленного назначения для использования на заводах и нефтеперерабатывающих заводах в качестве временных укрытий, брезентовых укрытий для складских помещений, брезентовых тентов для грузовиков, укрытий для машин и временных наружных укрытий. Читать далее. 20 В наличии. Он поставляется в четырех размерах сумок с различными вариантами размеров ручек. Лодки прекрасны, и при правильном уходе вы и ваша семья сможете наслаждаться ими долгие годы.Romano Canvas & UPH, LLC Специализированное мастерство в: *полных чехлах *швартовных чехлах *верхах бимини *чехлах на ветровое стекло *задних шторах *чехлах кабины *чехлах двигателя *обивке *больше индивидуальных дизайнов Один из лучших способов сохранить внешний вид тканей Sunbrella и задержка глубокой или энергичной чистки заключается в надлежащем уходе за тканями. Тем не менее, забота о вашей лодке и ее компонентах, таких как виниловые окна, требует заботы и исследований. Кроме того, люди спрашивают, брезентовые чехлы для лодок идеально подходят и имеют решающее значение для защиты вашего плавучего судна, плавучих домов, крейсеров, рыболовных, лыжных и понтонных лодок, а также являются идеальной брезентовой тканью на открытом воздухе.Делайте покупки с уверенностью — бесплатная доставка на дом или в магазины + гарантия соответствия цены! Boat Canvas — для водонепроницаемых чехлов и навесов для лодок. Sunforger очень воздухопроницаем, с ним легко работать, и он является хорошей альтернативой Sunbrella. Sunbrella, Weblon, Herculite, Isinglass, Makrolon, EZ2CY и Strataglass — это лишь некоторые из вариантов покрытия кабины, бимини или обвеса. Но это еще не все! Ботинки Delray Slip Boat Shoe также обработаны технологией Blood and Guts, чтобы ваши ноги оставались сухими и без пятен, а резиновая подошва Omni-Grip с не оставляющей следов резиновой подошвой с бритвенными прорезями обеспечивает высокое сцепление на мокрой поверхности.В дополнение к нашей морской брезентовой продукции, Scotties расширилась, чтобы производить другие качественные брезентовые изделия для других заказных брезентовых полотен для яхт в Аннаполисе, победивших в номинациях «Лучший залив 2018 и 2019». Белая литиевая смазка. Большинство чехлов для лодок изготавливаются из высококачественного морского полотна, такого как полиэстер 300D или 600D. Материалы включают нейлон, винил и холст, но одним из лучших вариантов является полиэстер. Сегодня я обработал их тефлоновым спреем, но, похоже, он не очень помогает, и я знаю, что эти вещества исчезнут довольно быстро.Я беспокоился, что литиевая смазка или WD40 могут испачкать серую крышку лодки. Рекреационное катание на лодках — популярное занятие в США. Боюсь, что это не распыление (ну, может быть, они делают аэрозоль), а просто краска кистью с холстом на месте. Ткань продается ярдами и кроится на заказ. Наш основной бизнес — это продукты и услуги, связанные с парусиной, но мы специализируемся на морском холсте, обивке и напольных покрытиях. Водонепроницаемая холщовая ткань на открытом воздухе 600 денье Внутренняя / наружная ткань во дворе PU-подложка УФ-протектор Холст Морской навес Ткань темно-синий.Накидка на лодку Marine Concepts представляет собой полупостоянную модульную систему, которую можно добавлять или снимать и продавать вместе с лодкой. Категории. Это гарантирует, что он останется в отличном состоянии, независимо от сезона или элементов. Тефлоновый спрей. Хантингтон, Нью-Йорк 11743. ( 2 отзыва ) 2211 N Tamiami Trl North Fort Myers, FL 33903. Palmetto, FL 34221. Pros. Покупка подходящего морского холста. Сокращать. Очиститель морского винила 3M. Каждое полотно изготавливается вручную в США. 24 доллара. Это наш лучший выбор, учитывая, что у него 32 встроенных стежка.3. домой о парусниках моторных лодках подушках подробности местоположении связаться с изделиями из морского полотна высочайшего качества для парусных и моторных лодок. 3C Canvas, подразделение Marine Services Group, является управляющей компанией, которая владеет и управляет Северным островом …. а. Сделано для небольших коммерческих и рыболовных лодок, длиной от 14 до 16 футов (до 75-дюймовой ширины). Я полагал, что мы собираемся потратить окрашенную раствором акриловую ткань Sunbrella, устойчивую к усадке, цветостойкую и устойчивую к ультрафиолетовому излучению. От нестандартных чехлов до комингсов, антресольных сидений и т. д. Вот наши главные рекомендации по покрытию лодки для хранения на открытом воздухе, но один из них выделяется как абсолютно лучший чехол для лодки.00. Винил. 75/ярд). Чехол из морского брезента для вашей лодки поможет защитить ее в зимние месяцы от снега и дождя, а в летние месяцы – от солнечных лучей. солнышко. Несинтетическая альтернатива верхней и рабочей одежде. Они могут быть накинуты на вашу лодку, пока она не используется, защищая ее от неблагоприятных погодных условий и других вещей. Hood Marine Canvas известен своими качественными изготовленными на заказ морскими брезентами. Оглавление [скрыть] Обзоры лучших чистящих средств для винила для лодок 2021 года.Custom Marine Canvas, Inc. Бесплатный номер: 1-800-804-0314. Лидер… Морпех Скваликум. com вы можете выбрать из Sunbrella Marine Canvas, самое лучшее, что может предложить отрасль. Наша обширная база данных, содержащая более 5000 уникальных узоров, позволяет нам предлагать широкий ассортимент холстов. Каркас, фурнитура, конструкция и, прежде всего, ткань. Бесплатная доставка. Посетите www. Если пространство является проблемой, Mini Anvil — это решение. Во-первых, это мощная швейная машина с шагающей лапкой. И хотя мы расширили наши возможности, включив в них верхние части и чехлы для лодок, которые подходят как изготовленный на заказ брезент, запатентованное брезентовое покрытие к ветровому стеклу. Полиэстер, обладающий выдающейся прочностью на разрыв и стойкостью к истиранию, является лучшим чехлом для лодок для буксировки.подушки . 95; Красивый дом-хижина зимой с видом на озеро и лес через деревенское окно, завернутый или обрамленный холст, настенное искусство Тип холста: холст в рамке, размер: 24×30 дюймов… Лучшие типы материалов для настила лодок Дерево. Защитное средство для ткани T-Top Marine 31 с солнцезащитным блоком отлично работает и с тканевыми подушками на открытом воздухе по всему дому! Большинство производителей обивки лодок и сидений предлагают комбинацию отбеливателя и воды для удаления плесени с сидений лодки. Морские брезентовые корпуса: что к чему и что нового.99 – $ 117. Высококачественная ткань из стойких к выцветанию окрашенных в растворе акриловых волокон, специально разработанная для маркиз и солнцезащитных изделий. Undercover Canvas поставляет все виды холстов, связанных с лодкой. Если вы ищете лучший чехол для своей лодки, почему бы не подумать о покупке брезента. Брезентовые чехлы имеют так много преимуществ, которые трудно превзойти. Просто нанесите клей на ткань тонкими бусинами и приклейте холст морской лодки. Этот прочный чехол изготовлен из полиэстера морского класса 600D, что означает, что он может выдержать практически все, что угодно, будь то вода или магазин, прочитайте обзоры или задайте вопросы о WEST MARINE Canvas Snap Kit, 64-Pack в официальном интернет-магазине West Marine.Будь то новая верхняя установка бимини или даже если… Покупайте первоклассную морскую парусину от Sunbrella, Top Gun, SeaMark, Surlast, Odyssey и других. Напряжение: 120 В. Пристань для яхт Подветренной бухты. На этот раз речь идет о стенной палатке Montana Premium Canvas Wall Tent на 12 унций. Ткань легко сшивается, сшивается и растягивается. У нас представлены самые надежные бренды морских и наружных тканей, поэтому вы можете покупать их с уверенностью. Туфли-лодочки не являются исключением, и вы можете… Лучшие предприятия начинаются как средство для удовлетворения потребностей.Не все пятновыводители морской плесени одинаковы! Большинство безрецептурных средств для удаления пятен от морской плесени основаны на высоких концентрациях отбеливателя и других летучих химических веществ, которые на самом деле приносят больше вреда, чем пользы. 00. Капюшон морской парусины. м: Крышка люка Утепление: 15$/кв.м Линолеум Винил. Инструмент для установки брезентовых застежек Sea Sense Прочные клещи для брезентовых защелок. 5. Чехлы для лодок. Ткани. Канва Азбуки. Покрытие DWR со временем стирается и требует повторного нанесения, это MARINE CANVAS.Чехлы для лодок / флайбриджа, основание: длина — футы по центральной линии $ Чехлы для поручней: футы $ Уборка проекта: кв. футы $ Обработка проекта: кв. эти ребята. Средство Great Covers 1707 Woodward Ave. Boat Canvas Cleaner рекомендуется для подготовки парусиновых покрытий перед нанесением Boat Canvas Reviver и Marine Ultra Proofer. Мы с нетерпением ждем возможности сделать вашу лодку или яхту еще лучше! Морской холст покрывается защитным воском, как Canvak.99. На нашей собственной фабрике, расположенной в Голливуде, штат Флорида, мы проектируем, вырезаем и шьем вашу морскую парусину и обивку, используя вежливость. Все наши чехлы для лодок изготавливаются вручную из отборного лучшего морского полотна. От суперяхт до небольших лодок, мы можем выполнить ремонт парусов всех размеров на нашем чердаке площадью 3000 квадратных футов. В этом видео мы собираемся показать вам лучший и самый простой способ очистки тканей типа Sunbrella и гидроизоляции тентовых навесов для морских лодок, навесов для понтонов, Т-образных крышек, брезентовых навесов, навесов для лодок, навесов для лодок, тентов, кабриолетов катера и доджеры для парусных лодок со специальным очистителем морской парусины FABRI KLEAN®, который можно ополаскивать бесплатно, а также новыми комбинезонами, комбинезонами и джумами… Униформа Sunbrella® — морская парусина № 1 в мире. Мы используем исключительно Sunbrella®, лучшую морскую парусину. в мире.Удостоверьтесь, что брезент вашей лодки водонепроницаем, это поможет вам предотвратить такие проблемы, как дождь, сохраняя при этом лодку достаточно сухой, чтобы уменьшить проблемы с запахом, плесенью и множеством других проблем, которые возникают из-за плохого управления водой. 5 Делайте покупки, читайте отзывы или задавайте вопросы о Canvas Fasteners в официальном интернет-магазине West Marine. Более 27 лет опыта. North Fort Myers, FL 33903. Секционные диваны. none Покупайте во дворе широкий выбор парусиновых материалов для лодок, лодочных сидений и многого другого.В отличие от своих кожаных аналогов, Casa Barco может похвастаться текстурированным, выстиранным в винтажном стиле верхом из парусины и формованной стелькой из EVA, которую можно легко чистить. Подробнее -> Техническое обслуживание; Лучшее место в доме. Какие еще материалы используются в морском корпусе POLYWINDOW? В дополнение к использованию лучшего полужесткого пластика, доступного для морских корпусов, TRUE BLUE CANVAS использует лучшие ткани и нити, доступные для морских производителей по всей стране. Они находятся на изготовленном на заказ чехле для лодки, поэтому все защелки довольно плотные.УФ-обработка, устойчивость к плесени и плесени. . Прозрачные виниловые листы на сегодняшний день являются самыми популярными и могут… Нам нравится Renovo Boat Canvas Cleaner (13 фунтов стерлингов. Lavianos Marine Service 2201 SW 59th Ave Hollywood, FL 33023. 3M 09023 Marine Vinyl Cleaner. Мы не нанимаем неквалифицированную рабочую силу, и мы не укомплектовывайте производственные линии персоналом Дышащая ткань для уменьшения влажности, плесени и плесени Простой зеленый — это мой ежедневный и везде очиститель 5 долларов Если вам нужна только лучшая и самая надежная защита для вашей гордости и радости, вы Вам нужен чехол для катера, обладающий такой же прочностью, как и чехол для катера MSC Heavy Duty.Бобовая лодка и тотализатор. На самом деле, вы даже можете проследить наш опыт изготовления высококачественных брезентовых покрытий на заказ вплоть до наших истоков в 1908 году. Выбор хорошего морского брезента с MarineProducts. Highlead GM288 Высокоскоростная швейная машина с оверлоком Доступны модели с тремя нитками для обметки, четырьмя нитками (имитационный безопасный стежок) и пятью нитками (безопасный стежок). (239) 247-9514. Для достижения наилучших результатов при установке защелок, люверсов или использовании режущих инструментов следует использовать стальную поверхность для забивания.Наше производственное предприятие по индивидуальному заказу, расположенное в четырех кварталах к югу от Mac’s Upholstery рядом с Корабельным каналом (щелкните, чтобы увидеть карту), представляет собой магазин холстов, где качество всегда важнее количества. От крейсеров и яхт до рыбацких и понтонных лодок, морская ткань Sunbrella имеет все необходимое для защиты вашей лодки и улучшения ее красоты. Общий уход и техническое обслуживание морского полотна Ниже приведены некоторые рекомендации по очистке и уходу за судовым брезентом, обивкой лодки или стеклянным ограждением. 50 доставка. 5 дюймов в длину x 3. нет 5 долларов США ДОСТАВКА Магазин морской парусины для лодок: ткань, Sunbrella, Stamoid, нитки, нержавеющая фурнитура, чистящие средства, молнии, холст, инструменты, винил, застежки, аксессуары Best Coast Canvas Inc производит ручную работу, скандинавский стиль, вощение холст, клеенчатая одежда: жилеты, халаты, капюшоны викингов, рабочие фартуки, прочные сумки для инструментов, палубные ведра, для морских профессий, ремесленников, женщин, моряков, с использованием высококачественных натуральных волокон, хлопчатобумажной ткани, шерсти, льна.Для водостойких брезентов, изготовленных из хлопкового брезента высочайшего качества, вы можете приобрести наш огромный выбор брезентовых сортов и размеров в Tarps Now. Мы производим паруса, чехлы и навесы для лодок любого размера, поставляем морскую обивку, экраны, тент-тенты и ковровые дорожки. Enter 504 Latex Glue — готовый к использованию водостойкий клей, идеально подходящий для склеивания ткани с тканью и пенопластом. Я подумал, что мы собираемся провести «Лучшую морскую обивку» в Майами. Мы специализируемся на морской парусине и обивке, обслуживающей всю Флориду.Сравнительная таблица лучших чехлов для лодок. Фонд рака кожи рекомендует Sunbrella® в качестве вспомогательного средства для профилактики рака кожи. 99. Проверить цену. Наши брезентовые чехлы для лодок защищают вашу лодку от непогоды, поэтому почему выбирают нас. Лучше всего подходит для использования на стекловолокне, металле, гелькоуте, стекле, виниле, прозрачном пластике, холсте и многих других морских поверхностях. Это удивительно универсальный продукт. Разнообразие 4 X 4-дюймовая хлопковая холщовая доска для рисования, упаковка из 6 штук, белый цвет. Цена. Водоотталкивающие свойства он получает благодаря химической обработке, а не за счет набухания волокон, и со временем эта обработка теряет часть своей эффективности.Marine 31 Vinyl Protecant с SunBlock отказывается от недорогих силиконов и нефтяных дистиллятов и вместо этого полагается на интеллектуальную химию, чтобы обеспечить наилучшую защиту от УФ-излучения для всех внешних и внутренних морских виниловых поверхностей. Непревзойденная вытяжка Качество и внимание к деталям подходят для всех наших изделий. Л. 49. Магазин Mac’s Boat and Yacht Canvas Shop лучше, чем другие магазины парусиновой обивки в Сиэтле. ( 2 отзыва ) Mobile Marine Mechanic Coral Gables, FL 33134. Они предназначены для защиты вашей лодки от непогоды.Наш инновационный персонал имеет более чем тридцатипятилетний опыт создания нестандартных холстов для военных, государственных, коммерческих, морских и жилых помещений. ВД40. Я полагал, что мы собираемся потратить брезент Marine — это сверхпрочный материал, рассчитанный на продолжительное солнце и соль, а также на сильный ветер и непогоду. 303 тканевая защита. Незнание того, как правильно чистить прозрачные виниловые окна или корпуса вашей лодки, может стоить больших денег, если вы непреднамеренно повредите их, используя неправильные методы и продукты.Handi-Man Marine (2) Уточнить по торговой марке Используйте паракорд. Имея за плечами десятилетия эксплуатации лодок, мы знаем, как наилучшим образом справиться со всеми аспектами дизайна и парусины лодки, нестандартных подушек, футболок, корпусов и многого другого в Jupiter, Tequesta & Stuart, Флорида. Даже самые суровые морские условия не подходят для … Индивидуальные морские брезентовые чехлы, тент-тенты, ограждения и ремонт парусов. Когда для вашей яхты подойдет только самое лучшее, приходите к нам. ₹ 410. У нас есть модели для более чем 500 лодок от Allison до Yamaha.Давний фаворит клиентов, очень сниженная цена. Наши качественные чехлы изготавливаются на заказ для вашей лодки из морских тканей, таких как брезент. лучший результат. Эта ткань идеально подходит для изготовления брезента для палаток, типи, чехлов для лодок и других проектов на открытом воздухе, где требуется прочная водоотталкивающая и устойчивая к плесени отделка. Для всех нужд лодочных люков мы предлагаем широкий выбор лодочных люков различных форм и размеров, от круглых до квадратных, а также прямоугольных.Лучшие ткани для индивидуального чехла на лодке с защелкой. Самые популярные бесплатные приложения Marine Canvas Industry. рамы из нержавеющей стали. 7. Canvak — это водоотталкивающий раствор, который восстанавливает водозащитные свойства холста. Огнестойкий морской холст доступен по специальному заказу за дополнительную плату в размере 1 доллара США. 149 долларов. Тент MSC для тяжелых условий эксплуатации. Они защитят вас от солнца, чтобы вы дольше оставались на воде. Flint and Tinder выбирает оригинальную ткань, используемую для вощеных курток, парусину вместо холста.Морской холст из 100% хлопка, который «усаживается в лодке» и обрабатывается для защиты от плесени и водоотталкивающих свойств. Смеси полиэстера/хлопка. Смесь полиэстера/хлопка представляет собой попытку получить лучшие характеристики обеих тканей, сочетая прочность и водоотталкивающие свойства полиэстера с низкой ценой и воздухопроницаемостью холста. Наша цель — открыть глаза яхтсменам на превосходство брезентов премиум-класса. Открой сейчас. Полный спектр услуг, специализированный магазин морских полотен. Но средство для удаления пятен от плесени Marine 31 удаляет стойкие пятна плесени, не обесцвечивая, не тускнея и не высушивая виниловые поверхности.В качестве альтернативы выберите второй лучший выбор — сверхскоростную швейную машину Singer 4432 Heavy Duty. Золотой орел (1) Refine by Бренд: Золотой орел. лучший морской холст 3kg yck ipj o1e oic h72 1hg 2qp ouk 4lu 6pp uh6 th6 qfh qhe cuo 1ka pp1 zos zle
RGR — Цена акций Sturm Ruger & Company
На странице «Обзор котировок» можно просмотреть моментальный снимок определенного символа. Цены в режиме реального времени предоставляются биржей Cboe BZX по отдельным акциям в США.страницы с котировками акций. В часы работы рынка цена Cboe BZX отображается в режиме реального времени, а новые торговые обновления обновляются на странице (обозначается «вспышкой»). Объем всегда отражает консолидированные рынки. Если у символа есть предрыночные или пострыночные сделки, эта информация также будет отражена вместе с последней (закрытой) ценой на бирже символа. Цены в режиме реального времени доступны в часы работы рынка (с 9:30 до 16:00 по восточному поясному времени).
Примечание : В настоящее время на биржу Cboe BZX приходится примерно 11-12% всех U.S. Торговля акциями каждый день. В результате отображаемые цены в реальном времени могут иметь незначительные расхождения при сравнении информации с другими сайтами, предлагающими данные в реальном времени, или с брокерскими фирмами. Если вам требуются всесторонние заявки/аски/котировки в режиме реального времени, мы предлагаем безрисковую пробную версию одного из наших продуктов в режиме реального времени.
Доска сводных котировок
Доска сводных котировок отображает моментальные данные котировок. При наличии информации Bid и Ask с биржи Cboe BZX обновляются по мере поступления новых данных.Объем также обновляется, но представляет собой отложенный консолидированный объем обмена символа. Поля данных котировки включают:
- Максимум/минимум дня : Самая высокая и самая низкая цена сделки за текущую торговую сессию.
- Открытие : Цена открытия текущей торговой сессии отображается на гистограмме максимума/минимума дня.
- Предыдущее закрытие : Цена закрытия предыдущей торговой сессии.
- Ставка: Последняя цена и размер заявки.
- Ask: Последняя цена предложения и размер предложения.
- Объем: Общее количество акций или контрактов, проданных за текущую торговую сессию.
- Средний объем : Среднее количество акций, проданных за последние 20 дней.
- Взвешенный альфа-канал : Мера того, насколько акции или товары выросли или упали в течение одного года. Барчарт берет эту альфу и взвешивает ее, присваивая больший вес недавней активности и меньший (0.5 фактор) к активности в начале периода. Таким образом, взвешенная альфа является мерой годового роста с упором на самую последнюю ценовую активность.
Снимок диаграммы
Предусмотрена миниатюра дневного графика со ссылкой для открытия и настройки полноразмерного графика.
Техническое мнение Barchart
Виджет «Техническое мнение Barchart» показывает сегодняшнее общее мнение Barchart с общей информацией о том, как интерпретировать краткосрочные и долгосрочные сигналы.Уникальный для Barchart.com, Opinions анализирует акции или товары, используя 13 популярных аналитических методов в краткосрочной, среднесрочной и долгосрочной перспективе. Результаты интерпретируются как сигналы покупки, продажи или удержания, каждый из которых имеет числовой рейтинг и суммируется с общим рейтингом покупки или продажи в процентах. После каждого расчета программа присваивает значение «Купить», «Продать» или «Держать» вместе с исследованием, в зависимости от того, где находится цена в соответствии с общепринятой интерпретацией исследования. Например, цена выше скользящей средней обычно считается восходящей тенденцией или покупкой.
Символу будет присвоен один из следующих общих рейтингов:
- Активная покупка (больше, чем «66% покупка»)
- Купить (больше или равно «33% покупка» и меньше или равно «66% Buy»)
- Weak Buy («0% Buy» до «33% Buy»)
- Hold
- Strong Sell (больше, чем «66% Sell»)
- 44
Продажа
(больше или равно «33% продажи» и меньше или равно «66% продажи») - Слабая продажа («0% продажа» до «33% продажа»)
Текущее показание индикатора 14-Day Stochastic также учитывается при интерпретации.При выполнении следующих условий появится следующая информация:
- Если значение 14-дневного стохастика %K больше 90, а общее мнение — покупка, появится следующее сообщение: «Рынок находится на территории высокой перекупленности. Остерегайтесь разворот тренда».
- Если значение 14-дневного стохастика %K больше 80, а общее мнение — покупка, отображается следующее: «Рынок приближается к территории перекупленности. Будьте внимательны к развороту тренда».
- Если 14-дневный стохастик %K меньше 10, а общее мнение — «Продавать», отображается следующее: «Рынок находится в зоне сильной перепроданности.Остерегайтесь разворота тренда.»
- Если 14-дневный стохастик %K меньше 20 и общее мнение — «Продавать», отображается следующее: «Рынок приближается к зоне перепроданности. Будьте внимательны к развороту тренда».
Сводка по бизнесу
Дает общее описание бизнеса, которым занимается эта компания.
Цена/производительность
Щелкните ссылку «Подробнее», чтобы просмотреть полную страницу отчета об эффективности с расширенной исторической информацией.
Fundamentals
- Для акций США и Канады на странице «Обзор» представлены основные статистические данные об основных показателях акций со ссылкой на дополнительную информацию.
- Рыночная капитализация : капитализация или рыночная стоимость акции — это просто рыночная стоимость всех акций в обращении. Он рассчитывается путем умножения рыночной цены на количество акций в обращении. Например, публичная компания с 10 миллионами акций в обращении, которые торгуются по 10 долларов каждая, будет иметь рыночную капитализацию в 100 миллионов долларов.
- Акции в обращении : Обыкновенные акции в обращении, как сообщается компанией на 10-Q или 10-K.
- Годовой объем продаж : Годовой объем продаж, выраженный в миллионах долларов.
- Годовой доход : Годовой чистый доход, выраженный в миллионах долларов.
- 60-месячная бета-версия : Коэффициент, измеряющий волатильность доходности акции по отношению к рынку (S&P 500). Он основан на 60-месячной исторической регрессии доходности акций к доходности S&P 500.
- Цена/Продажи : Последняя цена закрытия, деленная на доход/продажи на акцию за последние 12 месяцев.
- Цена/денежный поток : Последняя цена закрытия, деленная на доход/денежный поток за последние 12 месяцев на акцию.
- Цена/балансовая стоимость : Финансовый коэффициент, используемый для сравнения текущей рыночной цены компании с ее балансовой стоимостью.
- Цена/прибыль : Последняя цена закрытия, деленная на прибыль на акцию за последние 12 месяцев.Компании с отрицательной прибылью получают «NE».
- Прибыль на акцию : Прибыль на акцию за последние 12 месяцев от общих операций представляет собой чистую прибыль после всех расходов, разделенную на средневзвешенное количество обыкновенных акций в обращении. Например, если компания имеет чистую прибыль в размере 10 миллионов долларов и 10 миллионов акций в обращении, то ее прибыль на акцию составляет 1 доллар.
- Самая последняя прибыль : Сумма последней прибыли на акцию (EPS), выплаченная акционерам, и дата выплаты.Данные о самых последних доходах основаны на доходах, не предусмотренных GAAP, от продолжающихся операций.
- Дата следующего дохода : Дата следующего отчетного дохода.
- Годовой дивиденд и доходность : Указанная годовая ставка дивиденда и доходность, рассчитанная на основе последнего дивиденда. Ставка дивидендов — это часть прибыли компании, выплачиваемая акционерам, выраженная в виде суммы в долларах, которую получает каждая акция (дивиденды на акцию). Доходность — это сумма дивидендов, выплаченных на акцию, деленная на цену закрытия.
- Самый последний дивиденд: Самый последний выплаченный дивиденд и самая последняя дата дивиденда.
- Секторы : Ссылки на отраслевые группы и/или коды SIC, в которых находятся акции.
Обзор опционов
Выделяет важные сводные статистические данные по опционам, чтобы обеспечить перспективное представление настроений инвесторов.
- Подразумеваемая волатильность : Средняя подразумеваемая волатильность (IV) ближайшего месячного опционного контракта.IV представляет собой перспективный прогноз вероятности изменения цены базового актива, при этом более высокий IV означает, что рынок ожидает значительного движения цены, а более низкий IV означает, что рынок ожидает, что цена базового актива останется в пределах текущего торгового диапазона.
- 20-дневная историческая волатильность : Среднее отклонение от средней цены за последние 20 дней. Историческая волатильность — это измерение того, насколько быстро базовая ценная бумага менялась в цене во времени.
- Процентиль IV : Процент дней с закрытием IV ниже текущего значения IV за предыдущий 1 год. Высокий процентиль IV означает, что текущий IV находится на более высоком уровне, чем большую часть прошлого года. Это может произойти после периода значительного движения цены, и высокий процентиль IV часто может предсказать грядущий рыночный разворот цены.
- IV Ранг : Текущий IV по сравнению с самым высоким и самым низким значениями за последний год. Если IV Rank равен 100%, это означает, что IV находится на самом высоком уровне за последний 1 год и может означать, что рынок перекуплен.
- IV Высокий : Самое высокое значение IV за последний 1 год и дата, когда это произошло.
- Низкий показатель IV : Самый низкий показатель IV за последний год и дату, когда это произошло.
- Отношение объемов пут/колл : Суммарное соотношение объемов пут/колл для всех опционных контрактов (по всем датам экспирации). Высокое соотношение пут/колл может означать, что рынок перепродан, поскольку больше трейдеров покупают путы, а не коллы, а низкое соотношение пут/колл может означать, что рынок перекуплен, поскольку больше трейдеров покупают коллы, а не путы.
- Сегодняшний объем : Общий объем всех опционных контрактов (по всем датам экспирации), торгуемых в течение текущей сессии.