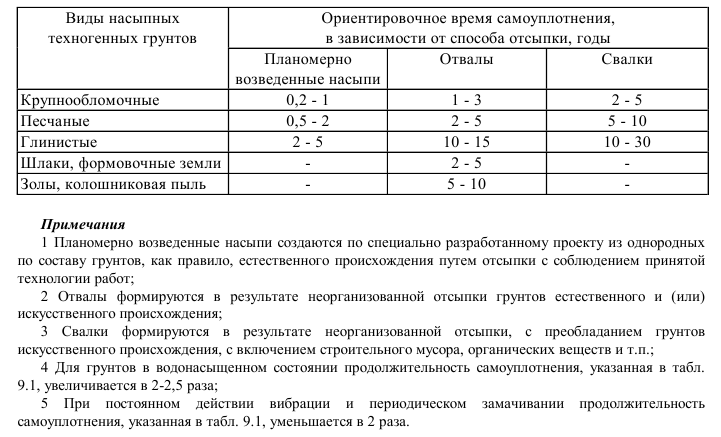
Уплотнение грунта СНиП 3.02.01-87
Строительству любых зданий или сооружений предшествует огромная работа по проектированию и подготовке застраиваемой площади к осуществлению планируемых мероприятий. Это относится ко всем объектам, которые планируется использовать длительный период, исчисляемый десятками и сотнями лет. Во время подготовки проводятся всевозможные пробы и тесты, показывающие пригодность площадей к дальнейшим действиям, например, берутся пробы грунта, вычисляется уровень подземных вод и прочие факторы. Если плотность грунта в зависимости от его типа не соответствует плановым показателям, проводится ряд мероприятий, направленных на уплотнение грунта.
Проведение подобных мероприятий способствует улучшению технических показателей, и как следствие решает ряд проблем, которые могут возникнуть в будущем, к примеру, проседание грунта со всеми вытекающими последствиями. Первым внешним проявлением проседания грунта могут стать трещины на стенах, а при наложении иных факторов к разрушению объекта.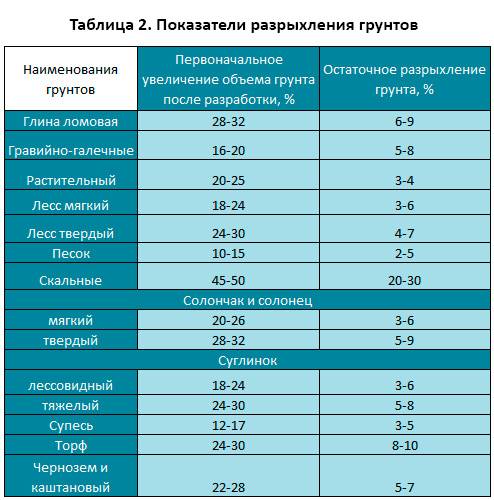
Методы уплотнения грунта
Качественный состав грунтов отличается в зависимости от географического расположения. При этом каждый из них имеет свою плотность, влажность и способность к проседанию. Поэтому для каждого вида почв разрабатывается комплекс мер, направленный на улучшение их характеристик, формирующий целую методологию.
Способность грунта к уплотнению определяется коэффициентом уплотнения, который вычисляется в лабораторных условиях соответствующими органами. И в зависимости от полученных показателей выбирается оптимальный метод уплотнения. При этом впоследствии рассчитывается усилие, прикладываемое для получения требуемого результата.
Условно методы разделяют на группы в зависимости от способа достижения цели – выведения воздуха из слоя почвы на заданной глубине. Так, различают поверхностные и глубинные способы. А категории оборудования и способы его применения выделяют статические, вибрационные, ударные и комбинированные методы, сочетающие в себе несколько видов влияния (давления).
Часть таких методов может применяться для малого частного строительства, другие же используются исключительно при возведении масштабных объектов при согласовании с местными властями, поскольку некоторые из них могут повлиять не только на заданную площадь, но и окружающие объекты и привести к их полному или частичному разрушению.
СНиП коэффициента уплотнения грунта
Все подобные операции четко регламентированы на законодательном уровне и поэтому их проведение ведется под контролем соответствующих организаций. Для того чтобы избежать возможных ошибок регламент и методология фиксируются в соответствующих документах – нормах (СНиП).
Уплотнение грунта и соответствующие показатели в российском законодательстве отражены в документах СП 45.13330.2012 и СНиП 3.02.01-87. Дата принятия этих документов отражена в их названии, соответственно первый принят в 2012 году, а второй в 1987, что означает его моральную изношенность.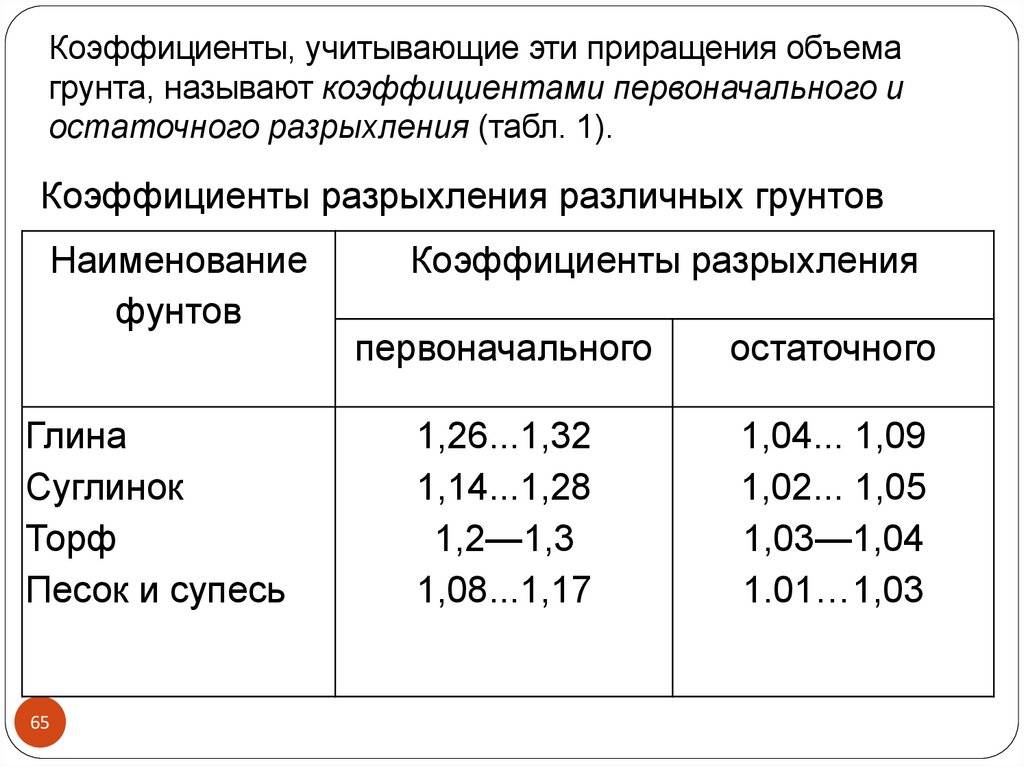 Однако действия, описанные в документах, актуализированы в 2013. Они описывают уплотнение грунтов разных видов и грунтовых подушек, укладываемых под фундамент сооружений различной конфигурации, в том числе подземной.
Однако действия, описанные в документах, актуализированы в 2013. Они описывают уплотнение грунтов разных видов и грунтовых подушек, укладываемых под фундамент сооружений различной конфигурации, в том числе подземной.
Каток для уплотнения грунта
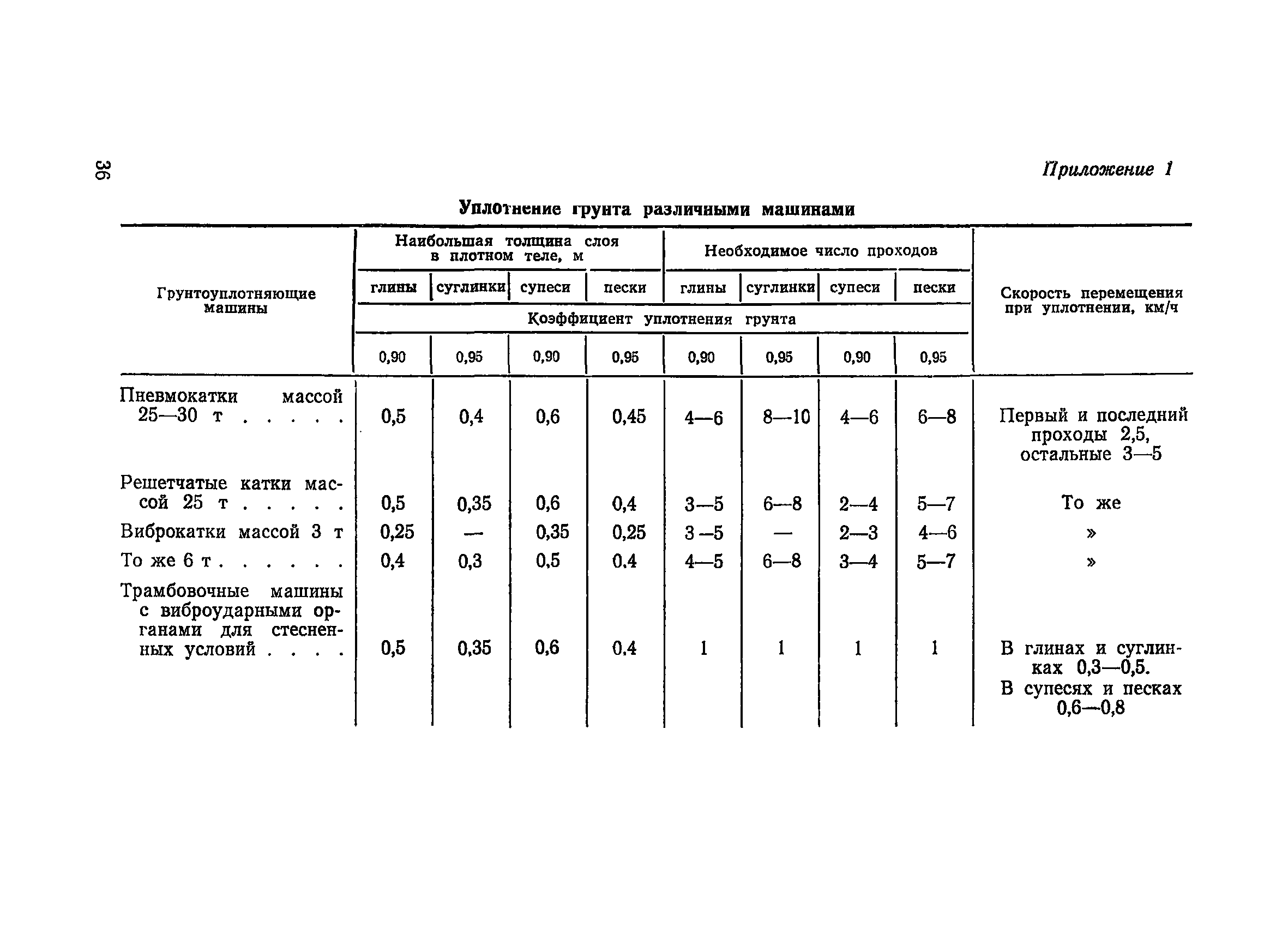
Для поверхностного уплотнения грунта применяются различного рода катки. В зависимости от характера работ и размеров строительного объекта они могут быть ручными и самоходными. Ручные обычно используются там, где сильное уплотнение почвы не требуется, например, при создании песчаной подушки для мощеных пешеходных дорожек. В данном конкретном случае сильных нагрузок вроде многотонной конструкции не будет, поэтому можно обойтись таким вариантом.
Самоходные и управляемые катки применяются на застраиваемых площадях круглой формы. Однако такой способ обработки не всегда приемлем, поэтому применяются катки, которые движутся не по спирали или кругу, а исключительно взад-вперед.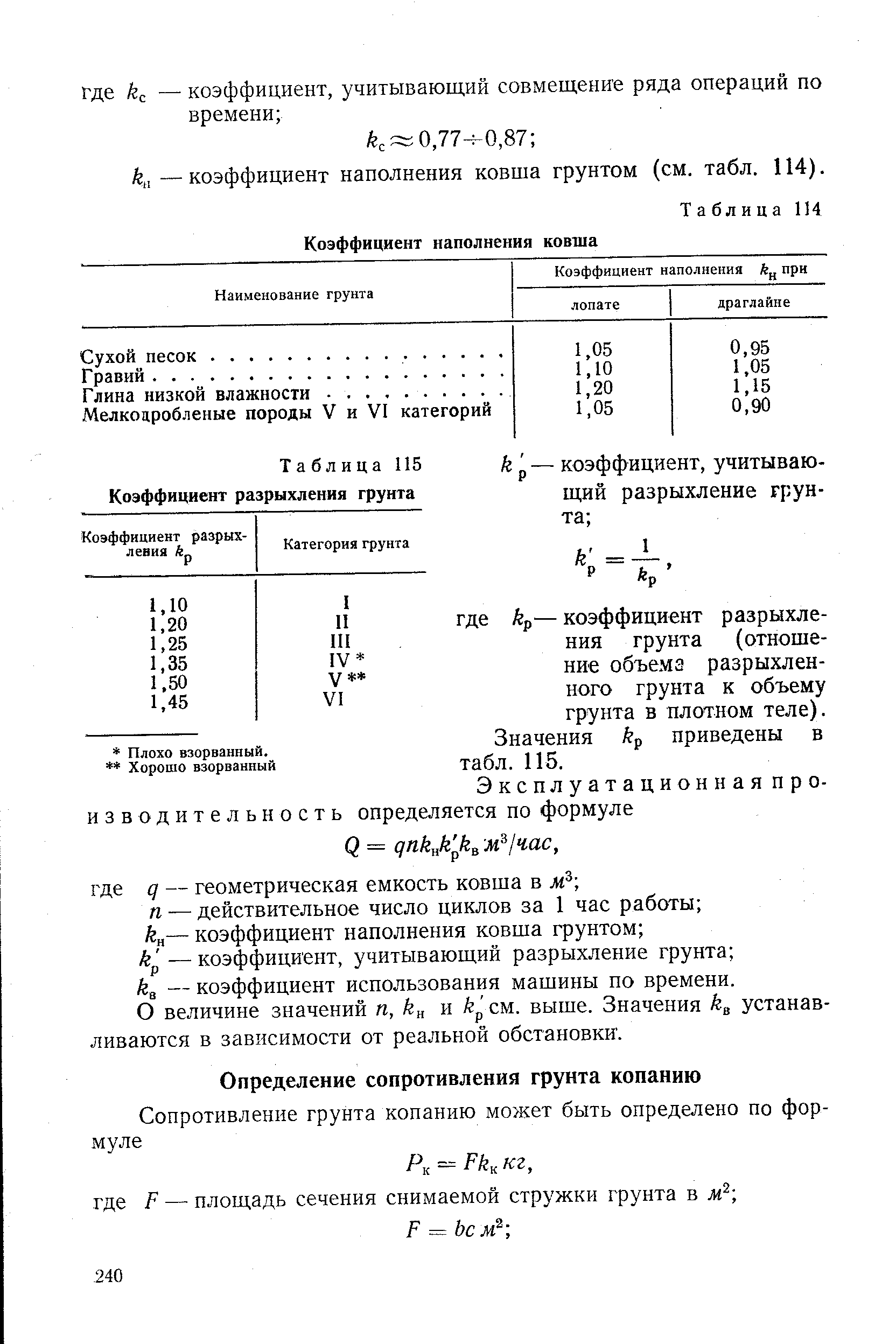
Катки для уплотнения грунта различаются по весу, одни из них по весу едва достигают в половину тонны, другие, напротив, имеют вес в десятки тонн.
Уплотнение грунта трамбовками
Определение коэффициента уплотнения грунта | Геологические изыскания
Результаты работ
Коэффициент уплотнения, полученный в ходе исследований, является основной для выявления несущей способности почвы.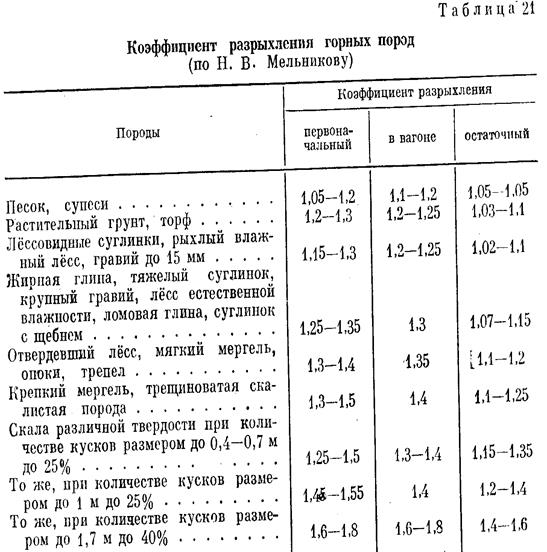 Таким образом, с помощью данного показателя производится оценка пригодности участка для возведения проектируемого сооружения. Полученный результат сравнивают с допустимыми нормативами и требованиями проекта.
Таким образом, с помощью данного показателя производится оценка пригодности участка для возведения проектируемого сооружения. Полученный результат сравнивают с допустимыми нормативами и требованиями проекта.
Важно знать!
Для масштабных проектов, которые оказывают существенную нагрузку на грунт, наряду с определением несущей способности, обязательно осуществляют расчет значений по предельным деформациям.Норма коэффициента уплотнения
Норма коэффициента уплотнения задается проектировщиками, в соответствии с задачами, целями и особенностями конкретного проекта. Задача изыскателей — определить, соответствуют ли фактические показатели заявленным требованиям.
Допустимые коэффициенты уплотнения почвы определяет нормативная база СНиП (пункты 3.02.01-87 и СП 45.13330.2012), обновленная в 2013-2014 гг.Здесь можно найти конкретные данные касательно допустимого уплотнения для определенных видов грунта и грунтовых подушек, которые используются при строительстве разных видов фундамента и строений, в том числе и подземных.
Коэффициент уплотнения варьируется в пределах от 0 до 1. Фактически он отражает уровень уплотненности почвы.
Для закладки основания бетонного ленточного фундамента нормой считается параметр уплотненности в >0,95 балла.
Стоимость работ
Наши эксперты проведут необходимые исследования и предоставят достоверные данные, которые исключат необходимость переделок на этапах проектирования и строительства.
Стоимость определения коэффициента уплотнения грунтового покрытия рассчитывается индивидуально в каждом конкретном случае.
Оставьте заявку на сайте или свяжитесь с нами любым удобным способом, чтобы получить бесплатную консультацию инженера-геолога. Мы оперативно рассчитаем стоимость проверки уплотнения почвы на вашем объекте.
при трамбовке, обратной засыпке, строительстве дороги
Что такое коэффициент уплотнения песка (Купл) знают не только специалисты, работающие в проектных организациях, но и эксплуатационники, основным видом деятельности которых является строительство. Его рассчитывают для того, чтобы сопоставить фактическую плотность на определенном участке, со значением, прописанном нормативных актах. Коэффициент уплотнения сыпучих материалов – это важный критерий, по которому оценивается качество выполнения подготовки к основным видам работ на строительных площадках.
Его рассчитывают для того, чтобы сопоставить фактическую плотность на определенном участке, со значением, прописанном нормативных актах. Коэффициент уплотнения сыпучих материалов – это важный критерий, по которому оценивается качество выполнения подготовки к основным видам работ на строительных площадках.
Что это такое?
Купл характеризует плотность, которую имеет грунт на конкретном участке, относится к тому же показателю материала, который перенес стандартное уплотнение в условиях лаборатории. Именно эта цифра применяется при оценке качества проведенных работ. Такой коэффициент определяет, насколько грунт на площадке соответствует требованиям ГОСТ 8736-93 и 25100-95.
При различных работах песок может иметь разный показатель плотности. Все эти нормы прописаны в СНиП 2.05.02-85, таблица 22. Еще их обычно указывают в проектных документах, в большинстве случаев этот показатель составляет от 0,95 до 0,98.
От чего изменяется коэффициент плотности
Если не понимать, что такое трамбовка песка, то посчитать правильно количество материала при строительстве практически не возможно. Ведь нужно четко знать, как повлияли на грунт различные манипуляции. То, какой коэффициент относительного уплотнения песка мы получим в конечном итоге, может зависеть от множества факторов:
- от способа перевозки;
- насколько длинным был маршрут;
- не появились ли повреждения механического характера;
- наличие посторонних вкраплений;
- попадание влаги.
Естественно, если вы заказали песок, то просто обязаны проверить его на месте, потому как поздние претензии будут совершенно неуместны.
Зачем учитывать относительный коэффициент при строительстве дорог
Этот показатель для песчаной подушки необходимо просчитать, и объясняется это обычным физическим явлением, которое знакомо любому человеку. Чтобы это понять, вспомните, как ведет себя взрыхленный грунт. Поначалу он рыхлый и объемный. Но уже спустя пару дней осядет и станет намного плотнее.
Чтобы это понять, вспомните, как ведет себя взрыхленный грунт. Поначалу он рыхлый и объемный. Но уже спустя пару дней осядет и станет намного плотнее.
Такая же участь ждет и любой другой сыпучий материал. Ведь его плотность увеличивается на складе под давлением собственного веса. Затем во время погрузки его взрыхляют, а уже непосредственно на стройплощадке опять происходит трамбовка песка своим весом. Кроме этого на грунт воздействует влажность. Песчаная подушка уплотнится при любых видах работ, будь то это строительство дорожного полотна, или обратная засыпка фундамента. Для всех этих факторов просчитаны соответствующие ГОСТ (8736-93 и 25100-95).
Как использовать относительный показатель
При любых строительных работах, одним из важнейших этапов считается составление сметы и подсчеты коэффициентов. Это нужно для того, чтобы правильно составить проект. Если важно узнать, как сильно уплотнится песок при транспортировке в самосвале или железнодорожном вагоне, достаточно найти в ГОСТ 8735-88 нужный показатель, и разделить на него требуемый объем.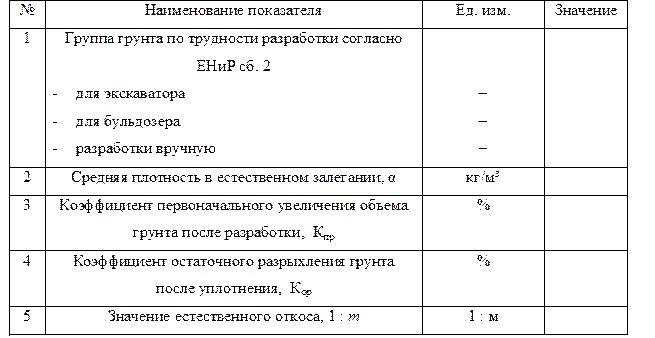
Необходимо учитывать и то, какие именно работы предстоят. То ли вы собираетесь делать песчаную подушку под дорожное полотно, или обратную засыпку фундамента. В каждой ситуации трамбовка будет проходить по-своему.
Например, при обратной засыпке песка наполняется вырытый котлован. Трамбовку делают при помощи различного оборудования. Иногда производят уплотнение виброплитой, но в некоторых случаях требуется каток. Соответственно и показатели будут разными. Учитывайте то, что грунт меняет свои свойства во время выемки. Так что количество засыпки нужно считать с учетом относительного показателя.
Таблица величин коэффициентов уплотнения в зависимости от назначения песка.
| Виды работ | Купл |
| Обратная засыпка котлованов | 0,95 |
| Обратная засыпка пазух | 0,98 |
| Обратная засыпка траншей | 0,98 |
| Восстановительный ремонт подземных инженерных сетей возле проезжей части дороги | 0,98 — 1 |
Пересеквенирование генома выявляет полиморфизм ДНК, связанный с определением размера / веса семян нута
Семена являются основным источником питания человека и животных. Нут — одна из коммерчески важных бобовых культур, обогащенных незаменимыми аминокислотами и ненасыщенными жирными кислотами [1]. Сообщалось о множестве генотипов нута с контрастирующими фенотипами, включая вариации размера / веса семян и качества семян [2,3]. Фенотипические вариации в основном объясняются естественным и искусственным отбором [4,5].Роль полиморфизма ДНК в управлении важными агрономическими признаками хорошо известна у сельскохозяйственных культур [[6], [7], [8]].
Нут — одна из коммерчески важных бобовых культур, обогащенных незаменимыми аминокислотами и ненасыщенными жирными кислотами [1]. Сообщалось о множестве генотипов нута с контрастирующими фенотипами, включая вариации размера / веса семян и качества семян [2,3]. Фенотипические вариации в основном объясняются естественным и искусственным отбором [4,5].Роль полиморфизма ДНК в управлении важными агрономическими признаками хорошо известна у сельскохозяйственных культур [[6], [7], [8]].
С появлением последовательности генома нута [2,9] исследования генотипирования и транскриптома ускорились беспрецедентными темпами. Было идентифицировано несколько простых повторов последовательности (SSR), чтобы использовать их в качестве маркеров для открытия генов-кандидатов, важных для агрономических признаков [10,11]. Разнообразные наборы генотипов нута были проанализированы, чтобы понять роль полиморфизмов ДНК в регулировании урожайности и качества семян [12,13].Однонуклеотидные полиморфизмы (SNP), связанные с размером / массой и качеством семян, были идентифицированы у нута с помощью полногеномных ассоциативных исследований [14,15]. Несколько локусов количественных признаков (QTL), связанных с размером / массой семян, были идентифицированы у нута с помощью различных подходов [12,13, [16], [17], [18], [19], [20]]. Эти исследования определили генетические маркеры с помощью повторного секвенирования генома, генотипирования путем секвенирования (GBS), масс-спектрофотометрии и / или гелевого анализа генотипов нута с контрастным весом семян / размера, которые наряду с фенотипическими данными использовались для идентификации QTL с использованием различных методы / программное обеспечение, такие как Cartographer, метод составного интервального отображения (CIM) и QTLNetwork [12,13, [16], [17], [18], [19], [20]].Более того, подход QTL-seq, который объединяет анализ групповой сегрегации на основе маркеров ДНК среди близкородственных генотипов и секвенирование следующего поколения, был использован для идентификации QTL, связанных с размером / массой семян у нута [12,18]. Например, две важные области генома (одна на хромосоме 4, а другая на хромосоме 1), несущие QTL, связанные с размером / массой семян, были идентифицированы с помощью QTL-seq [12].
Несколько локусов количественных признаков (QTL), связанных с размером / массой семян, были идентифицированы у нута с помощью различных подходов [12,13, [16], [17], [18], [19], [20]]. Эти исследования определили генетические маркеры с помощью повторного секвенирования генома, генотипирования путем секвенирования (GBS), масс-спектрофотометрии и / или гелевого анализа генотипов нута с контрастным весом семян / размера, которые наряду с фенотипическими данными использовались для идентификации QTL с использованием различных методы / программное обеспечение, такие как Cartographer, метод составного интервального отображения (CIM) и QTLNetwork [12,13, [16], [17], [18], [19], [20]].Более того, подход QTL-seq, который объединяет анализ групповой сегрегации на основе маркеров ДНК среди близкородственных генотипов и секвенирование следующего поколения, был использован для идентификации QTL, связанных с размером / массой семян у нута [12,18]. Например, две важные области генома (одна на хромосоме 4, а другая на хромосоме 1), несущие QTL, связанные с размером / массой семян, были идентифицированы с помощью QTL-seq [12].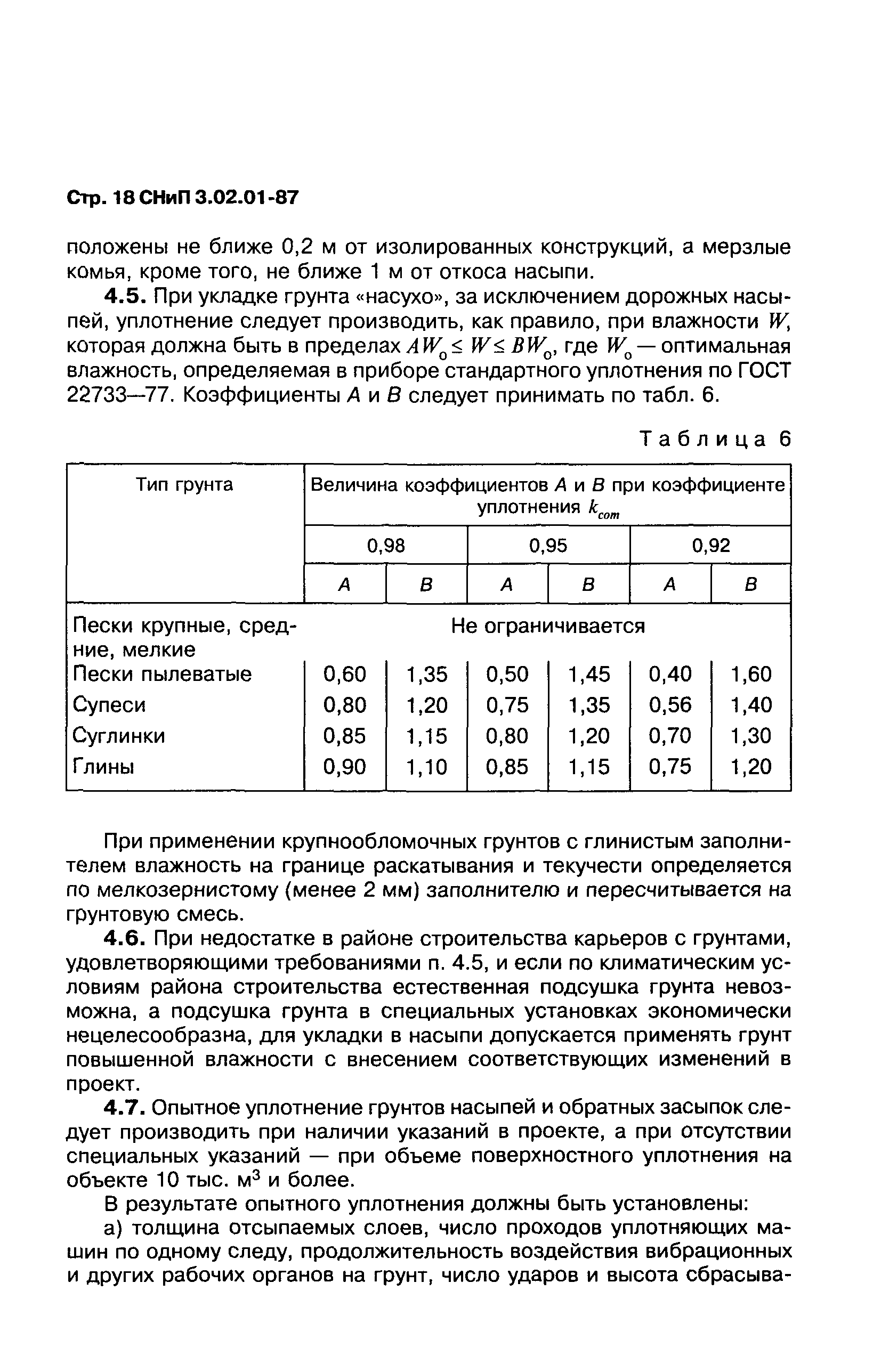 Аналогичным образом, другие исследования также выявили группы сцепления / геномные области / гены, которые могут определять размер / вес семян посредством анализа данных крупномасштабного генотипирования в многочисленных генотипах нута [12,13, [16], [17], [18], [18], [ 19], [20]].Недавно было обнаружено, что дифференциальная экспрессия генов, участвующих в росте / делении клеток и эндоредупликации между мелкосеменными и крупносемянными генотипами нута, определяет размер / вес семян нута [21]. Более того, предполагается важная роль факторов транскрипции (TF), расположенных внутри QTL, несущих полиморфизмы ДНК, в определении размера / веса семян [13,17,19].
Аналогичным образом, другие исследования также выявили группы сцепления / геномные области / гены, которые могут определять размер / вес семян посредством анализа данных крупномасштабного генотипирования в многочисленных генотипах нута [12,13, [16], [17], [18], [18], [ 19], [20]].Недавно было обнаружено, что дифференциальная экспрессия генов, участвующих в росте / делении клеток и эндоредупликации между мелкосеменными и крупносемянными генотипами нута, определяет размер / вес семян нута [21]. Более того, предполагается важная роль факторов транскрипции (TF), расположенных внутри QTL, несущих полиморфизмы ДНК, в определении размера / веса семян [13,17,19].
Генотипы нута классифицируются на типы Desi и kabuli на основании определенных характеристик, включая размер / вес семян и вариации цвета кожуры [2,22].В целом, семена генотипов нута дези меньше и темнее по цвету кожуры, чем семена нута кабули [23,24]. При филогенетическом древовидном анализе было обнаружено, что два типа нута сгруппированы вместе, что свидетельствует об их совместной эволюции [2]. В большинстве исследований генотипирования использовался метод GBS для уменьшения сложности генома и стоимости секвенирования для размещения большого количества образцов [16,17,25,26]. Однако GBS не позволяет анализировать весь геном, но больше подходит для анализа большой популяции и / или генотипов [26].Напротив, пересеквенирование всего генома (WGR) — это альтернативный метод анализа генотипов с высоким охватом и выявления большего количества и новых полиморфизмов ДНК, которые могут быть более актуальными для агрономических признаков и крупномасштабных приложений генотипирования [27,28] . Более того, SNP, обнаруженные с помощью WGR, часто используются в качестве эталона для проверки вызовов SNP, полученных с помощью подхода GBS, который включает в себя вменение недостающих данных из-за низкой глубины секвенирования и охвата, что указывает на полезность WGR для обнаружения полиморфизмов ДНК [29,30].
В большинстве исследований генотипирования использовался метод GBS для уменьшения сложности генома и стоимости секвенирования для размещения большого количества образцов [16,17,25,26]. Однако GBS не позволяет анализировать весь геном, но больше подходит для анализа большой популяции и / или генотипов [26].Напротив, пересеквенирование всего генома (WGR) — это альтернативный метод анализа генотипов с высоким охватом и выявления большего количества и новых полиморфизмов ДНК, которые могут быть более актуальными для агрономических признаков и крупномасштабных приложений генотипирования [27,28] . Более того, SNP, обнаруженные с помощью WGR, часто используются в качестве эталона для проверки вызовов SNP, полученных с помощью подхода GBS, который включает в себя вменение недостающих данных из-за низкой глубины секвенирования и охвата, что указывает на полезность WGR для обнаружения полиморфизмов ДНК [29,30].
Ранее мы проанализировали полиморфизмы ДНК между двумя мелкосеменными генотипами нута типа «кабули» и двумя крупносемянными генотипами нута кабульского типа с помощью ресеквенирования генома [31]. Это исследование выявило несколько генов-кандидатов, участвующих в определении размера / веса семян нута. Однако анализ большего набора генотипов нута с мелкими и крупными семенами, представляющих как желательные, так и кабули-типы, может дать лучшее понимание генов-кандидатов, участвующих в определении размера / веса семян. В этом исследовании мы проанализировали данные WGR 13 мелкосеменных и 5 крупносемянных генотипов нута желтого и кабульского типа для обнаружения важных полиморфизмов ДНК, которые могут определять размер / вес семян.Мы определили полиморфизмы ДНК, которые позволили дифференцировать мелкосемянные и крупнозернистые генотипы нута, и исследовали их правдоподобную функциональную значимость. Мы провели ассоциативный анализ, чтобы определить SNP, в значительной степени связанные с признаком размера / веса семян. Были идентифицированы гены-кандидаты, связанные с ростом / делением клеток, и гены, кодирующие TF, несущие в значительной степени ассоциированные SNP. Кроме того, выявлено влияние несинонимичных SNP на структуру и стабильность кодируемых белков.
Это исследование выявило несколько генов-кандидатов, участвующих в определении размера / веса семян нута. Однако анализ большего набора генотипов нута с мелкими и крупными семенами, представляющих как желательные, так и кабули-типы, может дать лучшее понимание генов-кандидатов, участвующих в определении размера / веса семян. В этом исследовании мы проанализировали данные WGR 13 мелкосеменных и 5 крупносемянных генотипов нута желтого и кабульского типа для обнаружения важных полиморфизмов ДНК, которые могут определять размер / вес семян.Мы определили полиморфизмы ДНК, которые позволили дифференцировать мелкосемянные и крупнозернистые генотипы нута, и исследовали их правдоподобную функциональную значимость. Мы провели ассоциативный анализ, чтобы определить SNP, в значительной степени связанные с признаком размера / веса семян. Были идентифицированы гены-кандидаты, связанные с ростом / делением клеток, и гены, кодирующие TF, несущие в значительной степени ассоциированные SNP. Кроме того, выявлено влияние несинонимичных SNP на структуру и стабильность кодируемых белков. В целом представленные здесь данные и результаты представляют собой ресурс для крупномасштабных приложений генотипирования и выявили гены-кандидаты, которые могут определять размер / вес семян нута.
В целом представленные здесь данные и результаты представляют собой ресурс для крупномасштабных приложений генотипирования и выявили гены-кандидаты, которые могут определять размер / вес семян нута.
Растения | Бесплатный полнотекстовый | Общегеномное исследование естественной изменчивости арабидопсиса, подверженного действию кислотных дренажных вод, и проверка ассоциированных генов с помощью обратной генетики
1. Введение
Загрязнение почвы является серьезной глобальной экологической проблемой, серьезно влияющей на производство сельскохозяйственных культур во всем мире, наряду с биоаккумуляцией. токсичных химикатов в нашей пище.Антропогенная деятельность является основным источником загрязнения почв [1]. Добыча угля является основной движущей силой изменений в землепользовании в регионе Аппалачи [2]. Промышленная добыча угля в Аппалачах ведется с середины 1800-х годов. Сегодня он продолжает вносить значительный вклад в региональную экономику и в энергетический портфель Администрации США [3]. Западная Вирджиния — второй по величине угледобывающий штат в США после Вайоминга (https: //www.eia.gov). Уголь добывается в Западной Вирджинии и других Аппалачских горах как подземными методами, так и методами удаления горных вершин. Горная добыча — это форма открытой добычи, которая включает удаление верхнего слоя почвы и обнажение угольных пластов. Обширные горные работы по добыче и переработке угля в Аппалачах и во всем мире оставили в наследство загрязненные горные породы. Сбросы из действующих или заброшенных шахт называются кислотными шахтными дренажами (AMD) и изменяют микробные сообщества и растительность [4,5 ].Вода AMD обычно кислая и содержит высокую концентрацию металлов, таких как кадмий (Cd), свинец (Pb), алюминий (Al), железо (Fe), селен (Se) и марганец (Mn). Эти значительные и неконтролируемые потоки элементов представляют собой серьезную угрозу как для экосистем, так и для здоровья человека в местной и региональной среде, окружающей рудники. Сульфиды металлов в окружающих породах и покрывающих породах окисляются под воздействием воздуха и воды, что приводит к образованию высококислотных продуктов выщелачивания [4], содержащих токсичные загрязнители [6].
Западная Вирджиния — второй по величине угледобывающий штат в США после Вайоминга (https: //www.eia.gov). Уголь добывается в Западной Вирджинии и других Аппалачских горах как подземными методами, так и методами удаления горных вершин. Горная добыча — это форма открытой добычи, которая включает удаление верхнего слоя почвы и обнажение угольных пластов. Обширные горные работы по добыче и переработке угля в Аппалачах и во всем мире оставили в наследство загрязненные горные породы. Сбросы из действующих или заброшенных шахт называются кислотными шахтными дренажами (AMD) и изменяют микробные сообщества и растительность [4,5 ].Вода AMD обычно кислая и содержит высокую концентрацию металлов, таких как кадмий (Cd), свинец (Pb), алюминий (Al), железо (Fe), селен (Se) и марганец (Mn). Эти значительные и неконтролируемые потоки элементов представляют собой серьезную угрозу как для экосистем, так и для здоровья человека в местной и региональной среде, окружающей рудники. Сульфиды металлов в окружающих породах и покрывающих породах окисляются под воздействием воздуха и воды, что приводит к образованию высококислотных продуктов выщелачивания [4], содержащих токсичные загрязнители [6]. Основные стрессы — дефицит фосфата (P), высокий уровень Al и протонная ризотоксичность (низкий pH) ограничивают рост растений в кислых почвах [7]. Кислотность угля AMD приводит к повышенной растворимости в почве тяжелых металлов, таких как Al, Fe и Mn, которые являются важными компонентами для роста и развития растений. Однако высокие концентрации этих ионов мешают критической ферментативной активности, что оказывает пагубное воздействие на растения [8]. Ионы водорода и катионы металлов заменяют основные катионы растений на участках катионного обмена, что приводит к вымыванию основных элементов из почвы [9].Избыточные концентрации также стимулируют образование свободных радикалов и активных форм кислорода (АФК), что приводит к окислительному стрессу и, как следствие, к повреждению клеток. Заброшенные шахты часто ускоряют процесс генерации AMD, а восстановление может потребовать десятилетий надлежащего управления. Сильно кислый pH (всего 2–4 стандартных единицы) в сочетании с токсичностью металлов может иметь серьезные последствия для водного биоразнообразия [10,11].
Основные стрессы — дефицит фосфата (P), высокий уровень Al и протонная ризотоксичность (низкий pH) ограничивают рост растений в кислых почвах [7]. Кислотность угля AMD приводит к повышенной растворимости в почве тяжелых металлов, таких как Al, Fe и Mn, которые являются важными компонентами для роста и развития растений. Однако высокие концентрации этих ионов мешают критической ферментативной активности, что оказывает пагубное воздействие на растения [8]. Ионы водорода и катионы металлов заменяют основные катионы растений на участках катионного обмена, что приводит к вымыванию основных элементов из почвы [9].Избыточные концентрации также стимулируют образование свободных радикалов и активных форм кислорода (АФК), что приводит к окислительному стрессу и, как следствие, к повреждению клеток. Заброшенные шахты часто ускоряют процесс генерации AMD, а восстановление может потребовать десятилетий надлежащего управления. Сильно кислый pH (всего 2–4 стандартных единицы) в сочетании с токсичностью металлов может иметь серьезные последствия для водного биоразнообразия [10,11]. У растений сложилась сложная сеть гомеостатических механизмов устойчивости к накоплению токсичных ионов.Эти механизмы включают снижение поглощения или оттока ионов металлов на плазматической мембране, хелатирование металлов в цитозоле и компартментализацию в вакуолях переносчиками, расположенными в тонопластах. Кроме того, восстановление поврежденных стрессом белков белком теплового шока или металлотионеином предотвращает накопление токсичных уровней ионов на чувствительных участках, помогая растениям переносить вредное воздействие ионов металлов [12]. У растений, которые могут расти на кислых почвах, развились механизмы, позволяющие выдерживать стресс ионной токсичности [13,14,15,16].Кроме того, растения в первую очередь реагируют на окислительное повреждение путем выработки АФК, таких как перекись водорода, свободные радикалы гидроксида и супероксид, которые влияют на физиологические и биохимические процессы растений [17]. Кислотность почвы также ослабляет опосредованное кальцием перекрестное сшивание пектина, участвующего в удлинении клеток, тем самым влияя на удлинение корня [9].
У растений сложилась сложная сеть гомеостатических механизмов устойчивости к накоплению токсичных ионов.Эти механизмы включают снижение поглощения или оттока ионов металлов на плазматической мембране, хелатирование металлов в цитозоле и компартментализацию в вакуолях переносчиками, расположенными в тонопластах. Кроме того, восстановление поврежденных стрессом белков белком теплового шока или металлотионеином предотвращает накопление токсичных уровней ионов на чувствительных участках, помогая растениям переносить вредное воздействие ионов металлов [12]. У растений, которые могут расти на кислых почвах, развились механизмы, позволяющие выдерживать стресс ионной токсичности [13,14,15,16].Кроме того, растения в первую очередь реагируют на окислительное повреждение путем выработки АФК, таких как перекись водорода, свободные радикалы гидроксида и супероксид, которые влияют на физиологические и биохимические процессы растений [17]. Кислотность почвы также ослабляет опосредованное кальцием перекрестное сшивание пектина, участвующего в удлинении клеток, тем самым влияя на удлинение корня [9]. При воздействии почвы на рудниках Аппалачей растения показали повышенную экспрессию переносчиков металлов, рецепторной киназы, хелатирующих белков, глутатиона. -s-трансфераза (GST), трехвалентная редуктаза, канальные белки и несколько генов сигнального пути [18].Это открытие предполагает, что несколько генов играют решающую роль в устойчивости растений к токсичности металлов. Следует отметить, что молекулярные механизмы, лежащие в основе токсичности металлов и устойчивости к кислотности, тесно взаимосвязаны. Подавление роста корней — наиболее частый эффект токсического действия ионов металлов на растения. Токсичность Al является одним из основных факторов, ограничивающих рост растений в кислых почвах, вызывая утолщение, укорочение и деформацию корней, влияя на деление клеток корня, тем самым влияя на удлинение корня [19,20,21].Кислотность может дополнительно нарушить сигнальные пути стресса, включая регуляцию активности свободного цитозоля Ca 2+ [22]. Многие виды лесов, инвазивных растений и пастбищ естественным образом адаптировались к кислым почвам, переносят H + и Al 3+, и процветают в почвах с pH 23].
При воздействии почвы на рудниках Аппалачей растения показали повышенную экспрессию переносчиков металлов, рецепторной киназы, хелатирующих белков, глутатиона. -s-трансфераза (GST), трехвалентная редуктаза, канальные белки и несколько генов сигнального пути [18].Это открытие предполагает, что несколько генов играют решающую роль в устойчивости растений к токсичности металлов. Следует отметить, что молекулярные механизмы, лежащие в основе токсичности металлов и устойчивости к кислотности, тесно взаимосвязаны. Подавление роста корней — наиболее частый эффект токсического действия ионов металлов на растения. Токсичность Al является одним из основных факторов, ограничивающих рост растений в кислых почвах, вызывая утолщение, укорочение и деформацию корней, влияя на деление клеток корня, тем самым влияя на удлинение корня [19,20,21].Кислотность может дополнительно нарушить сигнальные пути стресса, включая регуляцию активности свободного цитозоля Ca 2+ [22]. Многие виды лесов, инвазивных растений и пастбищ естественным образом адаптировались к кислым почвам, переносят H + и Al 3+, и процветают в почвах с pH 23]. Гены переносчиков органических кислот или анионов демонстрируют высокую экспрессию в проростках арабидопсиса, подвергнутых кислотному стрессу [24]. Кислотность почвы может повредить мембраны корней и активировать быстрый приток H + , деполяризуя плазматическую мембрану и приводя к подкислению цитоплазмы, что приводит к росту растений торможение [16,25,26].Растения, которые выживают при стрессе Al, по-видимому, стимулируют поглощение P, чтобы изменить баланс катион-анионов, обеспечивая выживание в токсичных Al-кислых почвах [27,28,29]. Обычными механизмами толерантности к Al является экссудация органической кислоты за счет активации анионных каналов и образования стабильных комплексов цитрата, оксалата и малата с Al для защиты верхушки корня [30,31,32]. Некоторые виды растений более устойчивы к Al, чем другие [33,34,35,36]. Однако молекулярные механизмы и связанные гены, которые придают толерантность к токсичности AMD, остаются малоизвестными.Арабидопсис, используемый в качестве модельного организма для понимания генетики и молекулярной биологии растений, имеет широкое естественное распространение в Европе, Азии и Северной Америке [37,38].
Гены переносчиков органических кислот или анионов демонстрируют высокую экспрессию в проростках арабидопсиса, подвергнутых кислотному стрессу [24]. Кислотность почвы может повредить мембраны корней и активировать быстрый приток H + , деполяризуя плазматическую мембрану и приводя к подкислению цитоплазмы, что приводит к росту растений торможение [16,25,26].Растения, которые выживают при стрессе Al, по-видимому, стимулируют поглощение P, чтобы изменить баланс катион-анионов, обеспечивая выживание в токсичных Al-кислых почвах [27,28,29]. Обычными механизмами толерантности к Al является экссудация органической кислоты за счет активации анионных каналов и образования стабильных комплексов цитрата, оксалата и малата с Al для защиты верхушки корня [30,31,32]. Некоторые виды растений более устойчивы к Al, чем другие [33,34,35,36]. Однако молекулярные механизмы и связанные гены, которые придают толерантность к токсичности AMD, остаются малоизвестными.Арабидопсис, используемый в качестве модельного организма для понимания генетики и молекулярной биологии растений, имеет широкое естественное распространение в Европе, Азии и Северной Америке [37,38]. Различия в естественных образцах Arabidopsis используются для раскрытия сложных генетических взаимодействий, лежащих в основе реакции растений на окружающую среду и эволюции морфологических признаков. Полногеномное ассоциативное исследование (GWAS) представляет собой многообещающий способ изучения сложных признаков, являющихся результатом множества генетических вариаций в широком спектре признаков растений.Однако исследования естественных вариаций адаптации к токсичности AMD по отношению к протонам и ионам металлов отсутствуют. В настоящем исследовании мы фенотипировали пять основных корневых признаков у 180 образцов Arabidopsis thaliana в ответ на условия роста с добавкой AMD. Эти данные были использованы в подходе GWAS для идентификации генов-кандидатов, вероятно, участвующих в адаптации к токсичности AMD.
Различия в естественных образцах Arabidopsis используются для раскрытия сложных генетических взаимодействий, лежащих в основе реакции растений на окружающую среду и эволюции морфологических признаков. Полногеномное ассоциативное исследование (GWAS) представляет собой многообещающий способ изучения сложных признаков, являющихся результатом множества генетических вариаций в широком спектре признаков растений.Однако исследования естественных вариаций адаптации к токсичности AMD по отношению к протонам и ионам металлов отсутствуют. В настоящем исследовании мы фенотипировали пять основных корневых признаков у 180 образцов Arabidopsis thaliana в ответ на условия роста с добавкой AMD. Эти данные были использованы в подходе GWAS для идентификации генов-кандидатов, вероятно, участвующих в адаптации к токсичности AMD.3. Обсуждение
В этом исследовании использовался GWAS для выявления геномных локусов у Arabidopsis, связанных с толерантностью к токсичности AMD, которая является глобальной проблемой в сельскохозяйственном производстве. Мы охарактеризовали пять основных фенотипов корней — совокупная длина корня, средний диаметр корня, площадь поверхности корня, объем корня и длина первичного корня — у 180 образцов Arabidopsis в ответ на ростовую среду с добавкой AMD. GWAS выявил гены, связанные с толерантностью к состояниям AMD. Большинство этих генов были факторами транскрипции, переносчиками анионов / катионов, переносчиками металлов и неизвестными белками. Два мутанта со вставкой Т-ДНК показали повышенную устойчивость к кислотности, особенно At1g63005 (miR399b) и At2g05635 (DEAD геликаза RAD3).Наши результаты GWAS и обратного генетического подхода предложили гены, устойчивые к угольной AMD.
Мы охарактеризовали пять основных фенотипов корней — совокупная длина корня, средний диаметр корня, площадь поверхности корня, объем корня и длина первичного корня — у 180 образцов Arabidopsis в ответ на ростовую среду с добавкой AMD. GWAS выявил гены, связанные с толерантностью к состояниям AMD. Большинство этих генов были факторами транскрипции, переносчиками анионов / катионов, переносчиками металлов и неизвестными белками. Два мутанта со вставкой Т-ДНК показали повышенную устойчивость к кислотности, особенно At1g63005 (miR399b) и At2g05635 (DEAD геликаза RAD3).Наши результаты GWAS и обратного генетического подхода предложили гены, устойчивые к угольной AMD.
 Низкий pH увеличивает растворимость металлов, таких как Al, Fe, Mn, Zn и Cu, и делает большие количества излишне доступными для растений в качестве токсичных питательных веществ [44].Когда pH почвы падает слишком низко, Al становится легко растворимым и оказывает вредное воздействие на сельскохозяйственные культуры [45]. Кислотность и токсичность Al являются преобладающими и связанными с ними токсичностями, поскольку Al растворяется только в кислой среде. Кислые почвы с низким pH могут привести к токсичности Al, Mn и Fe. Результаты многих исследований подчеркивают, что токсичность pH является важным ограничивающим фактором в кислой среде, особенно в очень кислых почвах. На практике сложно отделить эффекты кислотности как таковые от прямого воздействия повышенных концентраций токсичных катионов, таких как Al и Mn, в кислой среде.Образование богатого металлами кислотного раствора, известного как кислотный дренаж шахт (AMD), является серьезной экологической проблемой, связанной с горными работами.
Низкий pH увеличивает растворимость металлов, таких как Al, Fe, Mn, Zn и Cu, и делает большие количества излишне доступными для растений в качестве токсичных питательных веществ [44].Когда pH почвы падает слишком низко, Al становится легко растворимым и оказывает вредное воздействие на сельскохозяйственные культуры [45]. Кислотность и токсичность Al являются преобладающими и связанными с ними токсичностями, поскольку Al растворяется только в кислой среде. Кислые почвы с низким pH могут привести к токсичности Al, Mn и Fe. Результаты многих исследований подчеркивают, что токсичность pH является важным ограничивающим фактором в кислой среде, особенно в очень кислых почвах. На практике сложно отделить эффекты кислотности как таковые от прямого воздействия повышенных концентраций токсичных катионов, таких как Al и Mn, в кислой среде.Образование богатого металлами кислотного раствора, известного как кислотный дренаж шахт (AMD), является серьезной экологической проблемой, связанной с горными работами. Из-за воздействия AMD качество прилегающих поверхностных вод сильно ухудшается и в конечном итоге становится непригодным для поддержания биоразнообразия. Физико-химические свойства AMD от различных типов горных работ сильно различаются в зависимости от руды и металлов. Например, при добыче полезных ископаемых в Китае местные почвы были сильно загрязнены Cd, Cu и As, поэтому почва была непригодна для выращивания сельскохозяйственных культур [46].Результаты ICP – OES и физико-химические свойства показали, что AMD, полученная при добыче угля, имела значительно низкий pH и умеренные Al и Fe (Таблица 1). Поскольку низкие значения pH и содержания Al, Fe и Mn тесно взаимосвязаны в AMD, мы использовали AMD как единое целое, а не проводили эксперименты для каждого фактора в отдельности. Растения сидячие и не могут избежать кислотного стресса; однако большинство растений могут переносить умеренно кислую среду. Активность H + -АТФазы в плазматической мембране корневой клетки становится менее эффективной для вытеснения протонов в апопласт в кислых условиях.
Из-за воздействия AMD качество прилегающих поверхностных вод сильно ухудшается и в конечном итоге становится непригодным для поддержания биоразнообразия. Физико-химические свойства AMD от различных типов горных работ сильно различаются в зависимости от руды и металлов. Например, при добыче полезных ископаемых в Китае местные почвы были сильно загрязнены Cd, Cu и As, поэтому почва была непригодна для выращивания сельскохозяйственных культур [46].Результаты ICP – OES и физико-химические свойства показали, что AMD, полученная при добыче угля, имела значительно низкий pH и умеренные Al и Fe (Таблица 1). Поскольку низкие значения pH и содержания Al, Fe и Mn тесно взаимосвязаны в AMD, мы использовали AMD как единое целое, а не проводили эксперименты для каждого фактора в отдельности. Растения сидячие и не могут избежать кислотного стресса; однако большинство растений могут переносить умеренно кислую среду. Активность H + -АТФазы в плазматической мембране корневой клетки становится менее эффективной для вытеснения протонов в апопласт в кислых условиях. Однако и V-H + -АТФаза, и V-PPase перемещают избыточное количество H + из цитоплазмы в вакуоль [47] для поддержания pH цитоплазмы (7,0–7,5) [48]. Отток протонов регулирует концентрацию протонов в цитоплазме в вакуоль или апопластическое пространство с помощью протонных насосов [49,50,51]. Явление будет таким же — накопление или исключение — с более высокими концентрациями металлов. Механизмы устойчивости растений к стрессу Al включают исключение Al из кончиков корней и поглощение, но толерантность к Al в клетках корня.Естественно, некоторые виды растений разработали механизмы, снижающие кислотность, а другие нет, даже когда pH достигает всего 2,0. Некоторые виды растений, включая Acacia auriculiformis, выживают и образуют клубеньки при pH ≥ 2,0, а Acacia confusa и Melaleuca armillaris могут выжить при pH ≥ 2,5 [52]. Генотипы кукурузы демонстрируют генетическую изменчивость в отношении устойчивости к кислотности, что может быть использовано при разработке высокоурожайных генотипов кислотоустойчивой кукурузы [42].
Однако и V-H + -АТФаза, и V-PPase перемещают избыточное количество H + из цитоплазмы в вакуоль [47] для поддержания pH цитоплазмы (7,0–7,5) [48]. Отток протонов регулирует концентрацию протонов в цитоплазме в вакуоль или апопластическое пространство с помощью протонных насосов [49,50,51]. Явление будет таким же — накопление или исключение — с более высокими концентрациями металлов. Механизмы устойчивости растений к стрессу Al включают исключение Al из кончиков корней и поглощение, но толерантность к Al в клетках корня.Естественно, некоторые виды растений разработали механизмы, снижающие кислотность, а другие нет, даже когда pH достигает всего 2,0. Некоторые виды растений, включая Acacia auriculiformis, выживают и образуют клубеньки при pH ≥ 2,0, а Acacia confusa и Melaleuca armillaris могут выжить при pH ≥ 2,5 [52]. Генотипы кукурузы демонстрируют генетическую изменчивость в отношении устойчивости к кислотности, что может быть использовано при разработке высокоурожайных генотипов кислотоустойчивой кукурузы [42]. В поддержку этой концепции, около 30% из 30 генотипов риса верховых проявили устойчивость к кислотности почвы [53].В этих растениях нитрат (NTR1), ALMT1 и другие переносчики тяжелых металлов были активированы, чтобы выжить при ризотоксичности H + . Постоянное воздействие сильнокислых условий роста может существенно повлиять на структуру и функцию клеток корня [54]. В настоящем исследовании ~ 47% генов-кандидатов, идентифицированных в GWAS, участвовали в каталитических функциях. В общей сложности 43 из 737 генов-кандидатов были генами белков-переносчиков, что указывает на роль мембранного переносчика в клеточном гомеостазе. Эти гены включают АТФазы тяжелых металлов (HMA), NRAMP, семейство посредников диффузии катионов (CDF), семейство ZIP и катионные антипортерные белки [55,56,57].Кроме того, предполагается, что движение катионов металлов через мембраны будет вызвано движущей силой протонов, генерируемой насосами H + с питанием от АТФ. Кроме того, белки суперсемейства ABC являются первичными насосами, приводимыми в действие гидролизом АТФ и транспортируют широкий спектр субстратов, включая ионы, сахара, липиды, пептиды, пигменты, ксенобиотики и антибиотики.
В поддержку этой концепции, около 30% из 30 генотипов риса верховых проявили устойчивость к кислотности почвы [53].В этих растениях нитрат (NTR1), ALMT1 и другие переносчики тяжелых металлов были активированы, чтобы выжить при ризотоксичности H + . Постоянное воздействие сильнокислых условий роста может существенно повлиять на структуру и функцию клеток корня [54]. В настоящем исследовании ~ 47% генов-кандидатов, идентифицированных в GWAS, участвовали в каталитических функциях. В общей сложности 43 из 737 генов-кандидатов были генами белков-переносчиков, что указывает на роль мембранного переносчика в клеточном гомеостазе. Эти гены включают АТФазы тяжелых металлов (HMA), NRAMP, семейство посредников диффузии катионов (CDF), семейство ZIP и катионные антипортерные белки [55,56,57].Кроме того, предполагается, что движение катионов металлов через мембраны будет вызвано движущей силой протонов, генерируемой насосами H + с питанием от АТФ. Кроме того, белки суперсемейства ABC являются первичными насосами, приводимыми в действие гидролизом АТФ и транспортируют широкий спектр субстратов, включая ионы, сахара, липиды, пептиды, пигменты, ксенобиотики и антибиотики. Сетевой анализ наблюдателей взаимодействий с Arabidopsis подтвердил множество взаимодействий между партнерами из ядра, мембранных транспортеров, вакуолей и эндомембранной системы транспортировки.Факторы транскрипции являются критическими регуляторами во время стресса. Фактор транскрипции STOP1 регулирует AtALMT1 у арабидопсиса при низких значениях pH [58]. GWAS показал, что STOP1 связан с площадью поверхности корня, особенно с повышенной кислотностью. Проростки мутанта stop1 не смогли образовать корни в кислой среде AMD (данные не показаны), что указывает на то, что STOP1 контролирует обилие транскриптов набора генов, участвующих в толерантности к Al с низким pH. Al 3+ -индуцированная экспрессия плазматической мембраны AtALMT1 придает устойчивость к стрессу Al [23].Модификации клеточной стенки корня при токсичности Al играют важную роль в обеспечении устойчивости к стрессу Al [59]. НЕРЕГУЛЯРНАЯ КСИЛЕМА 11 (KNAT7) представляет собой репрессор транскрипции, который негативно регулирует транскрипцию для развития ксилемы и биогенеза вторичной клеточной стенки [60], а некоторые члены семейства — KNAT1, KNAT3, KNAT4 и KNAT5 — играют роль в регуляции развития корней [61].
Сетевой анализ наблюдателей взаимодействий с Arabidopsis подтвердил множество взаимодействий между партнерами из ядра, мембранных транспортеров, вакуолей и эндомембранной системы транспортировки.Факторы транскрипции являются критическими регуляторами во время стресса. Фактор транскрипции STOP1 регулирует AtALMT1 у арабидопсиса при низких значениях pH [58]. GWAS показал, что STOP1 связан с площадью поверхности корня, особенно с повышенной кислотностью. Проростки мутанта stop1 не смогли образовать корни в кислой среде AMD (данные не показаны), что указывает на то, что STOP1 контролирует обилие транскриптов набора генов, участвующих в толерантности к Al с низким pH. Al 3+ -индуцированная экспрессия плазматической мембраны AtALMT1 придает устойчивость к стрессу Al [23].Модификации клеточной стенки корня при токсичности Al играют важную роль в обеспечении устойчивости к стрессу Al [59]. НЕРЕГУЛЯРНАЯ КСИЛЕМА 11 (KNAT7) представляет собой репрессор транскрипции, который негативно регулирует транскрипцию для развития ксилемы и биогенеза вторичной клеточной стенки [60], а некоторые члены семейства — KNAT1, KNAT3, KNAT4 и KNAT5 — играют роль в регуляции развития корней [61]. Таким образом, идентификация гена семейства KNAT в нашем анализе не является сюрпризом. Модуляция углеводных полимеров в клеточной стенке корня может быть стратегией адаптации к избытку Al в кислых условиях роста.Показано, что такие модификации клеточной стенки ограничивают связывание токсичных ионов Al 3+ с клеточной стенкой [62]. Al, удерживаемый в клеточной стенке корня, транспортируется и секвестрируется в вакуоли корневых клеток для хранения [63]. Белок цинкового пальца имеет тенденцию локализоваться в мембране вакуоли и является неотъемлемым компонентом мембраны и выполняет такие функции, как связывание ионов цинка. Белки RING-finger C3HC4-типа были изучены в геномном масштабе у арабидопсиса и риса. Белки RING-finger арабидопсиса с предсказанными или известными биологическими функциями включают световую сигнализацию, секреторный путь, развитие корней [64] и устойчивость к стрессу [65].Идентификация этих генов в нашем анализе усиливает их функциональную роль в ответ на протонный и алюминиевый стресс при токсичности AMD.
Таким образом, идентификация гена семейства KNAT в нашем анализе не является сюрпризом. Модуляция углеводных полимеров в клеточной стенке корня может быть стратегией адаптации к избытку Al в кислых условиях роста.Показано, что такие модификации клеточной стенки ограничивают связывание токсичных ионов Al 3+ с клеточной стенкой [62]. Al, удерживаемый в клеточной стенке корня, транспортируется и секвестрируется в вакуоли корневых клеток для хранения [63]. Белок цинкового пальца имеет тенденцию локализоваться в мембране вакуоли и является неотъемлемым компонентом мембраны и выполняет такие функции, как связывание ионов цинка. Белки RING-finger C3HC4-типа были изучены в геномном масштабе у арабидопсиса и риса. Белки RING-finger арабидопсиса с предсказанными или известными биологическими функциями включают световую сигнализацию, секреторный путь, развитие корней [64] и устойчивость к стрессу [65].Идентификация этих генов в нашем анализе усиливает их функциональную роль в ответ на протонный и алюминиевый стресс при токсичности AMD. МикроРНК (миРНК) представляют собой класс коротких некодирующих РНК длиной около 22 нуклеотидов, которые играют ключевую роль в фундаментальных клеточных процессах, в том числе в том, как клетки реагируют на изменения окружающей среды или стрессы. Ген At1g63005 арабидопсиса кодирует реагирующую на фосфатное голодание miR399b, которая нацелена на PHO2, убиквитин-конъюгированный белок E2, который отрицательно влияет на содержание фосфата в побегах. Фосфор (P) дефицитен в большинстве кислых почв, поскольку растворимый неорганический P фиксируется Al и Fe [66].Мутант At1g63005 показал обширную корневую биомассу при обработке AMD. Однако гены-мишени miR399b, участвующие в придании толерантности к токсичности AMD и увеличению биомассы корней, все еще неизвестны. Недавно сверхэкспрессия miR169 (At3g26819) привела к повышенной засухоустойчивости томатов [67]. miR169 также участвует в регуляции образования клубеньков у клевера бочонка [68], подтверждая функции miRNA в компромиссе между защитой и ростом.
МикроРНК (миРНК) представляют собой класс коротких некодирующих РНК длиной около 22 нуклеотидов, которые играют ключевую роль в фундаментальных клеточных процессах, в том числе в том, как клетки реагируют на изменения окружающей среды или стрессы. Ген At1g63005 арабидопсиса кодирует реагирующую на фосфатное голодание miR399b, которая нацелена на PHO2, убиквитин-конъюгированный белок E2, который отрицательно влияет на содержание фосфата в побегах. Фосфор (P) дефицитен в большинстве кислых почв, поскольку растворимый неорганический P фиксируется Al и Fe [66].Мутант At1g63005 показал обширную корневую биомассу при обработке AMD. Однако гены-мишени miR399b, участвующие в придании толерантности к токсичности AMD и увеличению биомассы корней, все еще неизвестны. Недавно сверхэкспрессия miR169 (At3g26819) привела к повышенной засухоустойчивости томатов [67]. miR169 также участвует в регуляции образования клубеньков у клевера бочонка [68], подтверждая функции miRNA в компромиссе между защитой и ростом. Следовательно, идентификация генов-мишеней miR399b, связанных с устойчивостью к стрессу AMD и ростом корней, имеет важное значение.GWAS выявил несколько белков-переносчиков или мембранных белков, связанных с толерантностью к токсичности AMD. Транспортеры играют важную роль в защите растений от многих стрессов. Например, AT4G18790 кодирует природный белок макрофагов, связанный с устойчивостью (AtNRAMP5), который является переносчиком двухвалентных катионов для Fe 2+ и Mn 2+ . Недавнее исследование показало, что гомолог стрептококка NRAMP имеет решающее значение для выживания Streptococcus agalactiae в кислых условиях роста [69]. Эти белки, вероятно, локализуются на внутриклеточных мембранах, таких как пластидная оболочка и вакуолярная мембрана.У Arabidopsis избыточная экспрессия или нарушение генов NRAMP приводит к изменениям чувствительности к Fe или Cd. Существование семейства растительных NRAMP поднимает вопрос о важности динамической компартментализации металлов в растительных клетках [70].
Следовательно, идентификация генов-мишеней miR399b, связанных с устойчивостью к стрессу AMD и ростом корней, имеет важное значение.GWAS выявил несколько белков-переносчиков или мембранных белков, связанных с толерантностью к токсичности AMD. Транспортеры играют важную роль в защите растений от многих стрессов. Например, AT4G18790 кодирует природный белок макрофагов, связанный с устойчивостью (AtNRAMP5), который является переносчиком двухвалентных катионов для Fe 2+ и Mn 2+ . Недавнее исследование показало, что гомолог стрептококка NRAMP имеет решающее значение для выживания Streptococcus agalactiae в кислых условиях роста [69]. Эти белки, вероятно, локализуются на внутриклеточных мембранах, таких как пластидная оболочка и вакуолярная мембрана.У Arabidopsis избыточная экспрессия или нарушение генов NRAMP приводит к изменениям чувствительности к Fe или Cd. Существование семейства растительных NRAMP поднимает вопрос о важности динамической компартментализации металлов в растительных клетках [70]. Естественные вариации лежат в основе изменений экспрессии и функции транспортера алюминия NRAMP (NRAT1), которые играют решающую роль в толерантности риса к алюминию [71]. Как и AtALMT1 у Arabidopsis, GmALMT1 адаптирует сою к кислым почвам [72]. При высокой ризотоксичности H + цитоплазма корневых клеток начинает закисляться.Активируемая обоими транспортерами K + (AKT, HAK и CHX13), H + -АТФаза обеспечивает управляемую протонным насосом экструзию из цитоплазмы в апопласт. AtCHX13, протонный антипортер катионных мембран, расположен в плазматической мембране. AtCHX13 участвует в эндомембранной системе для регулирования pH [73]. БЕЛК 7 (PDLP7), расположенный в плазме, является неотъемлемым компонентом мембраны плазмодесм. PDLP7 играет роль в регулировании проницаемости плазмодесм для системного приобретенного иммунитета у растений [74].Функция PDLP7 может быть связана с перемещением H + внутрь и наружу клеток во время стресса. Прежде всего, мы обнаружили несколько переносчиков тяжелых металлов или членов семейства детоксикационных (At3g04900 и At4g16380), связанных с длиной первичного корня и площадью поверхности корня.
Естественные вариации лежат в основе изменений экспрессии и функции транспортера алюминия NRAMP (NRAT1), которые играют решающую роль в толерантности риса к алюминию [71]. Как и AtALMT1 у Arabidopsis, GmALMT1 адаптирует сою к кислым почвам [72]. При высокой ризотоксичности H + цитоплазма корневых клеток начинает закисляться.Активируемая обоими транспортерами K + (AKT, HAK и CHX13), H + -АТФаза обеспечивает управляемую протонным насосом экструзию из цитоплазмы в апопласт. AtCHX13, протонный антипортер катионных мембран, расположен в плазматической мембране. AtCHX13 участвует в эндомембранной системе для регулирования pH [73]. БЕЛК 7 (PDLP7), расположенный в плазме, является неотъемлемым компонентом мембраны плазмодесм. PDLP7 играет роль в регулировании проницаемости плазмодесм для системного приобретенного иммунитета у растений [74].Функция PDLP7 может быть связана с перемещением H + внутрь и наружу клеток во время стресса. Прежде всего, мы обнаружили несколько переносчиков тяжелых металлов или членов семейства детоксикационных (At3g04900 и At4g16380), связанных с длиной первичного корня и площадью поверхности корня. Гормональная регуляция модификации корня происходит во время кислотного стресса. Процессы закисления клеточной стенки в соответствии с теорией кислотного роста также могут происходить в корнях. Вызванное ауксином разрыхление и рост клеточных стенок в корнях катализируются группой ферментов, известных как экспансины, которые также связаны с реакцией растений на стресс с низким pH [75].Во время этой реакции рост и растяжимость клеточной стенки увеличиваются при низком pH. Сами корни могут влиять на pH ризосферы посредством притока или оттока катионов и анионов, активности H + -АТФазы, оттока органических кислот и дыхания корней. Другой гормональный регулятор, BREVIS RADIX (BRX, At1g31880), является позитивным регулятором передачи сигналов ауксина, необходимого для временного усиления полярного транспорта ауксина, необходимого для роста меристемы корня [76]. Внеклеточный pH влияет на активность ауксина и скорость пролиферации клеток в корневой меристеме у мутантов Arabidopsis.
Гормональная регуляция модификации корня происходит во время кислотного стресса. Процессы закисления клеточной стенки в соответствии с теорией кислотного роста также могут происходить в корнях. Вызванное ауксином разрыхление и рост клеточных стенок в корнях катализируются группой ферментов, известных как экспансины, которые также связаны с реакцией растений на стресс с низким pH [75].Во время этой реакции рост и растяжимость клеточной стенки увеличиваются при низком pH. Сами корни могут влиять на pH ризосферы посредством притока или оттока катионов и анионов, активности H + -АТФазы, оттока органических кислот и дыхания корней. Другой гормональный регулятор, BREVIS RADIX (BRX, At1g31880), является позитивным регулятором передачи сигналов ауксина, необходимого для временного усиления полярного транспорта ауксина, необходимого для роста меристемы корня [76]. Внеклеточный pH влияет на активность ауксина и скорость пролиферации клеток в корневой меристеме у мутантов Arabidopsis. Обнаружение гиперактивности протонных насосов в корнях brx может объяснить этот феномен и согласуется с устойчивым ростом и повышенной приспособленностью мутантов brx к чрезмерно кислой среде или почве. Новый аллель brx в образцах, недавно собранных из другого кислого участка отбора проб, демонстрирует аллели потери функции в природе и подтверждает их преимущество в кислых условиях pH почвы [77]. GWAS также выявил сильную ассоциацию с четырьмя ауксин-чувствительными белками семейства Gh4 на хромосоме 5, сильно связанными с диаметром корня.DWARF IN LIGHT 1 (DFL1, At5g54510), ауксин-чувствительный гомолог гена Gh4, отрицательно регулирует удлинение побеговых клеток и образование боковых корней [78]. Кроме того, индуцированный ауксином в культурах корней 12 (AIR12, At3g07390) рано экспрессируется во время индуцированного ауксином образования боковых корней у Arabidopsis [79]. GWAS также идентифицировал гены, связанные с брассиностероидами (BR), участвующие в толерантности к AMD. BR усиленная экспрессия 3 (BEE3), кодируемая At1g73830, представляет собой ДНК-связывающий регулятор транскрипции, активируемый BR.
Обнаружение гиперактивности протонных насосов в корнях brx может объяснить этот феномен и согласуется с устойчивым ростом и повышенной приспособленностью мутантов brx к чрезмерно кислой среде или почве. Новый аллель brx в образцах, недавно собранных из другого кислого участка отбора проб, демонстрирует аллели потери функции в природе и подтверждает их преимущество в кислых условиях pH почвы [77]. GWAS также выявил сильную ассоциацию с четырьмя ауксин-чувствительными белками семейства Gh4 на хромосоме 5, сильно связанными с диаметром корня.DWARF IN LIGHT 1 (DFL1, At5g54510), ауксин-чувствительный гомолог гена Gh4, отрицательно регулирует удлинение побеговых клеток и образование боковых корней [78]. Кроме того, индуцированный ауксином в культурах корней 12 (AIR12, At3g07390) рано экспрессируется во время индуцированного ауксином образования боковых корней у Arabidopsis [79]. GWAS также идентифицировал гены, связанные с брассиностероидами (BR), участвующие в толерантности к AMD. BR усиленная экспрессия 3 (BEE3), кодируемая At1g73830, представляет собой ДНК-связывающий регулятор транскрипции, активируемый BR. Гены BEE являются негативными регуляторами физиологических реакций на солевой стресс и засуху [80].Гормональная регуляция толерантности к низкому pH может быть важным механизмом у растений. Роль этих гормональных генов, связанных с AMD, требует дальнейшего изучения для понимания их роли в адаптации к стрессу при низком pH. Организация мембран и полярность клеток являются важным аспектом роста растений в условиях стресса. Эндомембранная система — это серия отсеков, которые упаковывают, маркируют и доставляют белки и молекулы. Частица транспортного белка (TRAPP) может играть жизненно важную роль в нацеливании и / или слиянии транспортных везикул эндоплазматического ретикулума-Гольджи с их акцепторным компартментом.TRAPP — это большой мультимерный белок, обладающий транспортной активностью. Транспортировка везикул контролирует структуры внутриклеточных компартментов и связь между клетками и окружающей средой для транспортировки белков. Сеть транс-Гольджи является основным центром секреторного и эндоцитарного транспорта для транспортировки клеточного груза к вакуолям через эндосомные сортировочные комплексы, необходимые для транспортного комплекса (ESCRT) [81].
Гены BEE являются негативными регуляторами физиологических реакций на солевой стресс и засуху [80].Гормональная регуляция толерантности к низкому pH может быть важным механизмом у растений. Роль этих гормональных генов, связанных с AMD, требует дальнейшего изучения для понимания их роли в адаптации к стрессу при низком pH. Организация мембран и полярность клеток являются важным аспектом роста растений в условиях стресса. Эндомембранная система — это серия отсеков, которые упаковывают, маркируют и доставляют белки и молекулы. Частица транспортного белка (TRAPP) может играть жизненно важную роль в нацеливании и / или слиянии транспортных везикул эндоплазматического ретикулума-Гольджи с их акцепторным компартментом.TRAPP — это большой мультимерный белок, обладающий транспортной активностью. Транспортировка везикул контролирует структуры внутриклеточных компартментов и связь между клетками и окружающей средой для транспортировки белков. Сеть транс-Гольджи является основным центром секреторного и эндоцитарного транспорта для транспортировки клеточного груза к вакуолям через эндосомные сортировочные комплексы, необходимые для транспортного комплекса (ESCRT) [81]. Вакуолярная сортировка белков 28 (VPS28) является частью комплекса ESCRT I. N-концевой домен гомологии эпсина (ENTH, At2g25430) играет важную роль в клатрин-опосредованном эндоцитозе в клетках корня [82].Эндоплазматический ретикулум — важная органелла, ответственная за синтез, сборку и сортировку секреторных белков. Транспорт белка между эндоплазматическим ретикулумом и аппаратом Гольджи опосредуется комплексами оболочки COPI и COPII. Sec23 / 24 (At4g14160) функционирует как часть COPII, опосредует транспорт белка [83]. Идентификация этих генов с помощью нашего анализа AMD GWAS открывает новые возможности для изучения важности клеточно-пузырькового транспорта и функций ESCRT для поддержания роста и развития растений в кислых условиях роста.ДНК-репарация DEAD-геликаза RAD3 может регулировать деление митотических клеток, УФ-ответ и окислительные стрессы, такие как толерантность к холоду [84,85]. Однако наш анализ не показал, что эта геликаза регулирует обильный рост корней в кислой среде.
Вакуолярная сортировка белков 28 (VPS28) является частью комплекса ESCRT I. N-концевой домен гомологии эпсина (ENTH, At2g25430) играет важную роль в клатрин-опосредованном эндоцитозе в клетках корня [82].Эндоплазматический ретикулум — важная органелла, ответственная за синтез, сборку и сортировку секреторных белков. Транспорт белка между эндоплазматическим ретикулумом и аппаратом Гольджи опосредуется комплексами оболочки COPI и COPII. Sec23 / 24 (At4g14160) функционирует как часть COPII, опосредует транспорт белка [83]. Идентификация этих генов с помощью нашего анализа AMD GWAS открывает новые возможности для изучения важности клеточно-пузырькового транспорта и функций ESCRT для поддержания роста и развития растений в кислых условиях роста.ДНК-репарация DEAD-геликаза RAD3 может регулировать деление митотических клеток, УФ-ответ и окислительные стрессы, такие как толерантность к холоду [84,85]. Однако наш анализ не показал, что эта геликаза регулирует обильный рост корней в кислой среде. Несколько белков LRR были связаны с корневыми признаками в нашем исследовании: белок семейства LRR (At4g13820), устойчивость к болезням TIR-NBS-LRR (At4g09430) и CC-NBS-LRR (At5g43740). Однако эти гены не связаны с толерантностью к кислоте. Как член кислых фосфатаз и расположенный во внеклеточной области, At3g52820 (ATPAP22) участвует в дефосфорилировании и связывании ионов металлов в кислой среде с высвобождением неорганического фосфата и играет ключевую роль. в метаболизме P [86].Другой член, BRG 1, выполняет молекулярную функцию активности убиквитин-протеинтрансферазы и связывания ионов металлов и связан с устойчивостью к болезни Botrytis cinerea [87]. Участие BRG1 в толерантности к AMD было бы интересной темой для исследования. В целом, наши результаты показывают, что гены-кандидаты, выбранные нами на основе GWAS, обладают большим потенциалом для использования в исследованиях ризотоксичности растений.
Несколько белков LRR были связаны с корневыми признаками в нашем исследовании: белок семейства LRR (At4g13820), устойчивость к болезням TIR-NBS-LRR (At4g09430) и CC-NBS-LRR (At5g43740). Однако эти гены не связаны с толерантностью к кислоте. Как член кислых фосфатаз и расположенный во внеклеточной области, At3g52820 (ATPAP22) участвует в дефосфорилировании и связывании ионов металлов в кислой среде с высвобождением неорганического фосфата и играет ключевую роль. в метаболизме P [86].Другой член, BRG 1, выполняет молекулярную функцию активности убиквитин-протеинтрансферазы и связывания ионов металлов и связан с устойчивостью к болезни Botrytis cinerea [87]. Участие BRG1 в толерантности к AMD было бы интересной темой для исследования. В целом, наши результаты показывают, что гены-кандидаты, выбранные нами на основе GWAS, обладают большим потенциалом для использования в исследованиях ризотоксичности растений.Дивергенция геномного острова приводит к эволюции меланизации корневого гриба галофита
Хоэкстра Х. Генетика, развитие и эволюция адаптивной пигментации у позвоночных. Наследственность. 2006; 97: 222–234.
CAS PubMed Google Scholar
Макнамара М.Э., Росси В., Слейтер Т.С., Роджерс С.С., Дюкрест А.Л., Дубей С. и др. Расшифровка эволюции меланина у позвоночных. Trends Ecol Evol. 2021; https://doi.org/10.1016/j.tree.2020.12.012.
Рулен А. Цветовой полиморфизм на основе меланина в ответ на изменение климата.Glob Chang Biol. 2014; 20: 3344–3350.
PubMed Google Scholar
Лоран С., Пфейфер С.П., Сеттлс М.Л., Хантер С.С., Хардвик К.М., Ормонд Л. и др. Популяционная геномика быстрой адаптации: разделение признаков отбора и демографии ящериц с белых песков. Mol Ecol. 2016; 25: 306–323.
CAS PubMed Google Scholar
Наранхо-Ортис, Массачусетс, Габальдон Т. Грибковая эволюция: основные экологические адаптации и эволюционные переходы. Биол Рев Камб Филос Соц. 2019; 94: 1443–1476.
Грибковая эволюция: основные экологические адаптации и эволюционные переходы. Биол Рев Камб Филос Соц. 2019; 94: 1443–1476.
PubMed PubMed Central Google Scholar
Кордеро Р.Дж., Касадевалл А. Функции грибкового меланина помимо вирулентности. Fungal Biol Rev.2017; 31: 99–112.
PubMed PubMed Central Google Scholar
Treseder KK, Lennon JT.Грибковые признаки, определяющие динамику экосистемы на суше. Microbiol Mol Biol Rev.2015; 79: 243–262.
CAS PubMed PubMed Central Google Scholar
Кейжар А., Гобец С., Племениташ А., Ленасси М. Меланин имеет решающее значение для роста черных дрожжей Hortaea werneckii в их естественной гиперсоленой среде. Fungal Biol. 2013; 117: 368–379.
PubMed Google Scholar

Сингаравелан Н., Гришкан И., Бехарав А., Вакамацу К., Ито С., Нево Э. Адаптивная реакция меланина почвенного гриба Aspergillus niger на стресс УФ-излучением в «Каньоне Эволюции», гора Кармель, Израиль. PLoS ONE. 2008; 3: e2993.
PubMed PubMed Central Google Scholar
Кришнан П., Мейле Л., Плиссонно С., Ма Х, Хартманн Ф. Э., Кролл Д. и др. Вставки мобильных элементов формируют регуляцию генов и выработку меланина у грибковых патогенов пшеницы.BMC Biol. 2018; 16:78.
PubMed PubMed Central Google Scholar
Перейра Д., Кролл Д., Бруннер П.С., Макдональд Б.А. Естественный отбор вызывает дивергенцию популяции для местной адаптации патогена пшеницы. Fungal Genet Biol. 2020; 141: 103398.
CAS PubMed Google Scholar
Desjardins CA, Giamberardino C, Sykes SM, Yu CH, Tenor JL, Chen Y, et al.
CAS PubMed PubMed Central Google Scholar
Робертсон К.Л., Мостагим А., Куомо К.А., Сото С.М., Лебедев Н., Бейли Р.Ф. и др. Адаптация черных дрожжей Wangiella dermatitidis к ионизирующему излучению: молекулярные и клеточные механизмы. PLoS ONE. 2012; 7: e48674.
CAS PubMed PubMed Central Google Scholar
Кнапп Д.Г., Немет Дж. Б., Барри К., Эно М., Хенриссат Б., Джонсон Дж. И др. Сравнительная геномика дает представление об образе жизни и выявляет функциональную неоднородность темных перегородчатых эндофитных грибов. Научный доклад 2018; 8: 6321.
PubMed PubMed Central Google Scholar
Фернандес Ч.В., Коидэ РТ.Функция меланина у эктомикоризного гриба Cenococcum geophilum в условиях водного стресса. Fungal Ecol. 2013; 6: 479–486.
Fungal Ecol. 2013; 6: 479–486.
Google Scholar
Редман Р.С., Шихан КБ, Стаут Р.Г., Родригес Р.Дж., Хенсон Дж.М. Термостойкость, вызванная симбиозом растений и грибов. Наука. 2002; 298: 1581.
CAS PubMed Google Scholar
Пи К.Г., Кеннеди П.Г., Талбот Дж. М..Размеры биоразнообразия в микобиоме Земли. Nat Rev Microbiol. 2016; 14: 434–447.
CAS PubMed Google Scholar
Родригес Р.Дж., Уайт Д.Ф., Арнольд А.Е., Редман Р.С. Грибковые эндофиты: разнообразие и функциональные роли. N Phytol. 2009. 182: 314–330.
CAS Google Scholar
Yuan ZL, Su ZZ, Zhang CL. Понимание биоразнообразия и функций эндофитов корневых грибов: аскомицет Harpophora oryzae как модельный случай.В кн .: Ирина С Дружинина И.С., ред. Кубичек С.П.). Mycota Vol. IV: взаимоотношения между окружающей средой и микробами. 3-е изд. Springer; 2016, стр 205–214.
Кубичек С.П.). Mycota Vol. IV: взаимоотношения между окружающей средой и микробами. 3-е изд. Springer; 2016, стр 205–214.
Бертело С., Лейвал С., Фулон Дж., Шало М., Блаудес Д. Стимуляция роста растений, выработка метаболитов и устойчивость к металлам темных септатных эндофитов, выделенных из загрязненных металлами участков фитоменеджмента тополя. FEMS Microbiol Ecol. 2016; 92: fiw144.
PubMed Google Scholar
Hill PW, Broughton R, Bougoure J, Havelange W, Newsham KK, Grant H и др. Симбиоз покрытосеменных с немикоризными грибами-партнерами усиливает усвоение азота из древнего органического вещества в теплеющей морской Антарктике. Ecol Lett. 2019; 22: 2111–2119.
PubMed PubMed Central Google Scholar
Матеу М., Болдуин А., Мол Дж., Ярвуд С. Эндофит с темной перегородкой улучшает солеустойчивость местных и инвазивных линий Phragmites australis . ISME J. 2020; 14: 1943–1954.
ISME J. 2020; 14: 1943–1954.
Google Scholar
Поррас-Альфаро А., Эррера Дж., Синсабо Р.Л., Оденбах К.Дж., Лоури Т., Натвиг Д.О. Новый консорциум корневых грибов, связанных с доминирующей пустынной травой. Appl Environ Microbiol. 2008. 74: 2805–2813.
CAS PubMed PubMed Central Google Scholar
Qin Y, Pan XY, Kubicek CP, Druzhinina IS, Chenthamara K, Labbé J, et al.Разнообразные плейоспоральные грибы, ассоциированные с растениями, из засоленных районов: экологическая устойчивость и влияние статуса азота на рост растений. Front Microbiol. 2017; 8: 158.
PubMed PubMed Central Google Scholar
Gostinčar C, Grube M, de Hoog S, Zalar P, Gunde-Cimerman N. Экстремотолерантность у грибов: эволюция на грани. FEMS Microbiol Ecol. 2010; 71: 2–11.
PubMed Google Scholar
Юань З.Л., Дружинина И.С., Лаббе Дж., Редман Р., Цинь Й., Родригес Р. и др. Специализированный микробиом галофита и его роль в обеспечении устойчивости растений, не являющихся хозяевами, к засолению. Научный доклад 2016; 6: 32467.
CAS PubMed PubMed Central Google Scholar
Yuan ZL, Druzhinina IS, Wang X, Zhang X, Peng L, Labbé J. Взгляд на высокополиморфный эндофит, выделенный из корней галофитной водоросли suaeda salsa : Laburnicola rhiz101 sp.ноя (Didymosphaeriaceae, Pleosporales). Fungal Biol. 2020; 124: 327–337.
CAS PubMed Google Scholar
Ellison CE, Hall C, Kowbel D, Welch J, Brem RB, Glass NL, et al. Популяционная геномика и локальная адаптация диких изолятов модельного микробного эукариота. Proc Natl Acad Sci USA. 2011; 108: 2831–2836.
CAS PubMed PubMed Central Google Scholar

Li H, Durbin R. Быстрое и точное выравнивание в режиме длительного чтения с преобразованием Барроуза – Уиллера. Биоинформатика 2010; 26: 589–595.
PubMed PubMed Central Google Scholar
Ли Х, Хандакер Б., Вайсокер А, Феннелл Т., Руан Дж., Гомер Н. и др. Формат выравнивания / карты последовательностей и SAMtools. Биоинформатика. 2009. 25: 2078–2079.
PubMed PubMed Central Google Scholar
Van der Auwera GA, Carneiro MO, Hartl C, Poplin R, Del Angel G, Levy-Moonshine A и др. От данных FastQ до вызовов вариантов с высокой степенью достоверности: конвейер лучших практик инструментария анализа генома. Curr Protoc Bioinform. 2013; 43: 11.10.1–11.10.33.
Google Scholar
Чанг С.К., Чоу С.К., Теллиер Л.К., Ваттикути С., Перселл С.М., Ли Дж.Дж. PLINK второго поколения: ответ на вызов более крупных и богатых наборов данных. Gigascience.2015; 4: 7.
Gigascience.2015; 4: 7.
PubMed PubMed Central Google Scholar
Ян Дж., Ли С.Х., Годдард М.Э., Вишер П.М. GCTA: инструмент для анализа сложных признаков в масштабе всего генома. Am J Hum Genet. 2011; 88: 76–82.
CAS PubMed PubMed Central Google Scholar
Александр Д.Х., Новембре Дж., Ланге К. Быстрая модельная оценка родословной неродственных особей.Genome Res. 2009; 19: 1655–1664.
CAS PubMed PubMed Central Google Scholar
Хусон Д.Х., Брайант Д. Применение филогенетических сетей в эволюционных исследованиях. Mol Biol Evol. 2006; 23: 254–267.
CAS PubMed PubMed Central Google Scholar
Wilken M, Steenkamp E, Wingfield M, De Beer ZW, Wingfield B. Какой ген MAT? Пересмотрена номенклатура генов брачного типа Pezizomycotina (Ascomycota). Fungal Biol Rev.2017; 31: 199–211.
Fungal Biol Rev.2017; 31: 199–211.
Google Scholar
Альтшул С.Ф., Мэдден Т.Л., Шеффер А.А., Чжан Дж., Чжан З., Миллер В. и др. Gapped BLAST и PSI-BLAST: новое поколение программ поиска по базам данных белков. Nucleic Acids Res. 1997; 25: 3389–3402.
CAS PubMed PubMed Central Google Scholar
Райт С. Генетическая структура популяций.Энн Ойген. 1951; 15: 323–354.
CAS PubMed Google Scholar
Nei M (ed). Молекулярно-эволюционная генетика. Издательство Колумбийского университета; 1987.
Карлсон С.С., Томас Д.Д., Эберле М.А., Суонсон Дж. Э., Ливингстон Р. Дж., Ридер М. Дж. И др. Геномные области, демонстрирующие положительный отбор, идентифицированные из плотных данных генотипа. Genome Res. 2005; 15: 1553–1565.
CAS PubMed PubMed Central Google Scholar

Pembleton LW, Cogan NOI, Forster JW. StAMPP: пакет R для расчета генетической дифференциации и структуры популяций смешанного уровня плоидности. Мол Экол Ресур. 2013; 13: 946–952.
CAS PubMed Google Scholar
Axelsson E, Ratnakumar A, Arendt ML, Maqbool K, Webster MT, Perloski M, et al. Геномная подпись приручения собаки свидетельствует об адаптации к диете, богатой крахмалом. Природа. 2013; 495: 360–364.
CAS PubMed Google Scholar
Юй Г, Ван LG, Хань И, Хэ Цюй. clusterProfiler: пакет R для сравнения биологических тем среди генных кластеров. Омикс. 2012; 16: 284–287.
CAS PubMed PubMed Central Google Scholar
Чжао С., Гиббонс Дж. Популяционная геномная характеристика вариации числа копий условно-патогенного грибка Aspergillus fumigatus . PLoS ONE. 2018; 13: e0201611.
2018; 13: e0201611.
PubMed PubMed Central Google Scholar
Klambauer G, Schwarzbauer K, Mayr A, Clevert DA, Mitterecker A, Bodenhofer U, et al. cn.MOPS: смесь Пуассона для обнаружения вариаций числа копий в данных секвенирования следующего поколения с низким уровнем ложного обнаружения. Nucleic Acids Res. 2012; 40: e69.
CAS PubMed PubMed Central Google Scholar
Таджима Ф. Статистический метод проверки гипотезы нейтральной мутации по полиморфизму ДНК. Генетика.1989; 123: 585–595.
CAS PubMed PubMed Central Google Scholar
Nei M, Li WH. Математическая модель для изучения генетической изменчивости с точки зрения эндонуклеаз рестрикции. Proc Natl Acad Sci USA. 1979; 76: 5269–5273.
CAS PubMed PubMed Central Google Scholar
Hutter S, Vilella AJ, Rozas J. Анализ полиморфизма ДНК по всему геному с использованием VariScan.BMC Bioinform. 2006; 7: 409.
Google Scholar
Вагнер Д.Н., Барис Т.З., Даян Д.И., Ду Х, Олексяк М.Ф., Кроуфорд Д.Л. Мелкомасштабная генетическая структура из-за адаптивной дивергенции между микропредприятиями. Наследственность. 2017; 118: 594–604.
CAS PubMed PubMed Central Google Scholar
Rech GE, Sanz-Martín JM, Anisimova M, Sukno SA, Thon MR. Естественный отбор по кодирующим и некодирующим последовательностям ДНК связан с генами вирулентности в патогенных грибах растений.Genome Biol Evol. 2014; 6: 2368–2379.
CAS PubMed PubMed Central Google Scholar
Стеркен Р., Кикенс Р., Коппенс Е., Веркаутерен И., Забо М., Инзе Д. и др. Исследование популяционной геномики основных генов клеточного цикла арабидопсиса показывает признаки естественного отбора. Растительная клетка. 2009; 21: 2987–2998.
Растительная клетка. 2009; 21: 2987–2998.
CAS PubMed PubMed Central Google Scholar
Yu F, Keinan A, Chen H, Ferland RJ, Hill RS, Mignault AA и др. Обнаружение естественного отбора путем эмпирического сравнения со случайными участками генома. Hum Mol Genet. 2009. 18: 4853–4867.
CAS PubMed PubMed Central Google Scholar
Nielsen R, Williamson S, Kim Y, Hubisz MJ, Clark AG, Bustamante C. Геномное сканирование для выборочного сканирования с использованием данных SNP. Genome Res. 2005; 15: 1566–1575.
CAS PubMed PubMed Central Google Scholar
Павлидис П., Живкович Д., Стаматакис А., Алачиотис Н. SweeD: вероятностное обнаружение выборочных разверток в тысячах геномов. Mol Biol Evol. 2013; 30: 2224–2234.
CAS PubMed PubMed Central Google Scholar

Барретт Дж. К., Фрай Б., Маллер Дж., Дейли М. Дж.. Haploview: анализ и визуализация карт LD и гаплотипов. Биоинформатика. 2005; 21: 263–265.
CAS Google Scholar
Zhan F, He Y, Zu Y, Li T, Zhao Z. Характеристика меланина, выделенного из темного перегородчатого эндофита (DSE), Exophiala pisciphila . Мир J Microbiol Biotechnol. 2011; 27: 2483–2489.
CAS Google Scholar
Тейлор Дж. У., Ханн-Соден С., Бранко С., Сильвен И., Эллисон К. Э.. Клональное размножение грибов. Proc Natl Acad Sci USA. 2015; 112: 8901–8908.
CAS PubMed PubMed Central Google Scholar
McGuire IC, Davis JE, Double ML, MacDonald WL, Rauscher JT, McCawley S и др. Образование гетерокарионов и парасексуальная рекомбинация между вегетативно несовместимыми линиями в популяции гриба каштанового ожога, Cryphonectria parasitica . Mol Ecol. 2005; 14: 3657–3669.
Mol Ecol. 2005; 14: 3657–3669.
CAS PubMed Google Scholar
Szulkin M, Gagnaire PA, Bierne N, Charmantier A. Популяционные геномные следы мелкомасштабной дифференциации между местообитаниями средиземноморских голубых синиц.Mol Ecol. 2016; 25: 542–558.
CAS PubMed Google Scholar
Hamilton JA, De la Torre AR, Aitken SN. Мелкомасштабные изменения окружающей среды вносят свой вклад в интрогрессию в трехвидовом гибридном комплексе ели. Древовидные генетические геномы. 2015; 11: 817.
Google Scholar
Йеман С. Геномные перестройки и эволюция кластеров локально адаптивных локусов.Proc Natl Acad Sci USA. 2013; 110: E1743 – E1751.
CAS PubMed PubMed Central Google Scholar
Крокен С., Гласс Н.Л., Тейлор Дж. В., Йодер О.К., Тюрджен Б.Г. Филогеномный анализ генов поликетидсинтазы I типа у патогенных и сапробных аскомицетов. Proc Natl Acad Sci USA. 2003; 100: 15670–15675.
В., Йодер О.К., Тюрджен Б.Г. Филогеномный анализ генов поликетидсинтазы I типа у патогенных и сапробных аскомицетов. Proc Natl Acad Sci USA. 2003; 100: 15670–15675.
CAS PubMed PubMed Central Google Scholar
Woo PCY, Tam EW, Chong KT, Cai JJ, Tung ET, Ngan AH и др. Высокое разнообразие генов поликетидсинтазы и кластера генов биосинтеза меланина в Penicillium marneffei . FEBS J. 2010; 277: 3750–3758.
CAS PubMed Google Scholar
Камеяма К., Монтегю П.М., Слушающий VJ. Экспрессия рецепторов меланоцит-стимулирующего гормона коррелирует с пигментацией млекопитающих и может модулироваться интерферонами.J. Cell Physiol. 1988. 137: 35–44.
CAS PubMed Google Scholar
Upadhyay S, Xu X, Lowry D, Jackson JC, Roberson RW, Lin X. Субклеточная компартментализация и перемещение биосинтетического аппарата для грибкового меланина. Cell Rep. 2016; 14: 2511–2518.
Cell Rep. 2016; 14: 2511–2518.
CAS PubMed PubMed Central Google Scholar
Коулман Дж. Дж., Милонакис Э. Отток грибов: la pièce de résistance.PLoS Pathog. 2009; 5: e1000486.
PubMed PubMed Central Google Scholar
Cosgrove DJ. Микробные экспансины. Annu Rev Microbiol. 2017; 71: 479–497.
CAS PubMed Google Scholar
Влияние внекорневой подкормки салициловой кислоты и оксида азота на уменьшение вызванного дефицитом железа хлороза Arachis hypogaea L. | Ботанические исследования
Bradford MM: Быстрый и чувствительный метод количественного определения количества белка в микрограммах, использующий принцип связывания белок-краситель. Annu Rev Biochem 1976, 72: 248–254. 10.1016 / 0003-2697 (76)
-3
CAS Статья Google Scholar
Брискин Д. П., Леонард Р.Т., Ходжес TK: Выделение плазматической мембраны: маркеры и общие принципы. Метод Энзимол 1987, 148: 542–558. DOI: 10.1016 / 0076–6879 (87) 48053–1 DOI: 10.1016 / 0076-6879 (87) 48053-1
П., Леонард Р.Т., Ходжес TK: Выделение плазматической мембраны: маркеры и общие принципы. Метод Энзимол 1987, 148: 542–558. DOI: 10.1016 / 0076–6879 (87) 48053–1 DOI: 10.1016 / 0076-6879 (87) 48053-1
CAS Статья Google Scholar
Бьюкенен ББ, Груиссем У., Джонс Р.Л .: Биохимия и молекулярная биология растений .Роквилл, Мэриленд: Американское общество физиологов растений; 2000.
Google Scholar
Буш DS: Регулирование цитозольного кальция у растений. Физиология растений 1993, 103: 7–13.
CAS PubMed PubMed Central Google Scholar
Cakmak I, Marschner H: Дефицит магния и высокая интенсивность света усиливают активность супероксиддисмутазы, аскорбатпероксидазы и глутатионредуктазы в листьях фасоли. Физиология растений 1992, 98: 1222–1227. 10.1104 / стр.98.4.1222
10.1104 / стр.98.4.1222
CAS Статья PubMed PubMed Central Google Scholar
Chen WW, Yang JL, Qin C, Jin CW, Mo JH, Ye T, Zheng SJ: Оксид азота действует после ауксина, вызывая активность хелатредуктазы корня железа в ответ на дефицит железа у арабидопсиса. Физиология растений 2010, 154: 810–819.DOI: 10.1104 / pp.110.161109 DOI: 10.1104 / pp.110.161109 10.1104 / pp.110.161109
CAS Статья PubMed PubMed Central Google Scholar
Ding F, Wang XF, Shi QH, Wang ML, Yang FJ, Gao QH: Экзогенный оксид азота смягчал ингибирование фотосинтеза и активности антиоксидантных ферментов в железодефицитной китайской капусте (Brassica chinensis L.). Agric Sci China 2008, 7: 168–179.10.1016 / S1671-2927 (08) 60036-X
CAS Статья Google Scholar
Ding H, Duan L, Wu H, Yang R, Ling H, Li WX, Zhang F: Регулирование AhFRO1, Fe (III) -хелатредуктазы арахиса, во время стресса, связанного с дефицитом железа, и при пересадке кукурузы.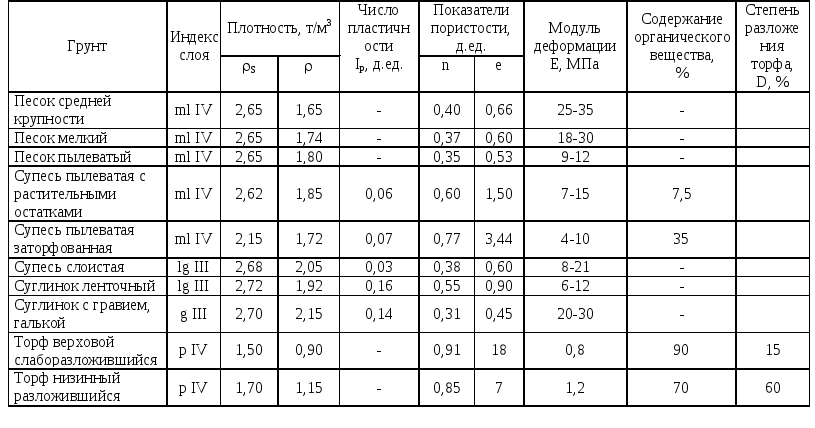 Physiol Plant 2009, 136: 274–283. DOI: 10.1111 / j.1399–3054.2009.01219.x DOI: 10.1111 / j.1399-3054.2009.01219.x 10.1111 / j.1399-3054.2009.01219.x
Physiol Plant 2009, 136: 274–283. DOI: 10.1111 / j.1399–3054.2009.01219.x DOI: 10.1111 / j.1399-3054.2009.01219.x 10.1111 / j.1399-3054.2009.01219.x
CAS Статья PubMed Google Scholar
Ding H, Duan LH, Li F, Yan HF, Zhao M, Zhang FS, Li WX: Клонирование и функциональный анализ транспортера железа арахиса AhIRT1 во время стресса от дефицита железа и скрещивания с кукурузой. J Plant Physiol 2010, 167: 996–1002. doi.org/10.1016/j.jplph.2009.12.019 doi.org/10.1016/j.jplph.2009.12.019 10.1016 / j.jplph.2009.12.019
CAS Статья PubMed Google Scholar
Эль-Тайеб МА: Реакция зерен ячменя на интерактивное воздействие солености и салициловой кислоты. Регулятор роста растений 2005, 45: 215–224. DOI: 10.1007 / s10725–005–4928–1 DOI: 10.1007 / s10725-005-4928-1 10.1007 / s10725-005-4928-1
CAS Статья Google Scholar
Эль-Тайеб М. А., Эль-Энани А.Е., Ахмед Н.Л .: Адаптивный ответ, вызванный салициловой кислотой, на медный стресс у подсолнечника (Helianthus annuus L.). Регулятор роста растений 2006, 50: 191–199. DOI: 10.1007 / s10725–006–9118–2 DOI: 10.1007 / s10725-006-9118-2 10.1007 / s10725-006-9118-2
А., Эль-Энани А.Е., Ахмед Н.Л .: Адаптивный ответ, вызванный салициловой кислотой, на медный стресс у подсолнечника (Helianthus annuus L.). Регулятор роста растений 2006, 50: 191–199. DOI: 10.1007 / s10725–006–9118–2 DOI: 10.1007 / s10725-006-9118-2 10.1007 / s10725-006-9118-2
CAS Статья Google Scholar
Farooq M, Basra SMA, Wahid A, Rehman H: Экзогенно применяемый оксид азота повышает устойчивость к засухе мелкозернистого ароматного риса (Oryza sativa L.). J Agronomy & Crop Science 2009, 195: 254–261. DOI: 10.1111 / j.1439–037X.2009.00367.x DOI: 10.1111 / j.1439-037X.2009.00367.x 10.1111 / j.1439-037X.2009.00367.x
CAS Статья Google Scholar
Frédéric G, Duby G, Stedingk EV, Zhao RM, Morsomme P, Boutry M: Экспрессия конститутивно активированной плазматической мембраны H + -ATPase изменяет развитие растений и повышает устойчивость к соли. Физиология растений 2007, 144: 1763–1776. DOI: 10.1104 / pp.107.103762 DOI: 10.1104 / pp.107.103762 10.1104 / pp.107.103762
Физиология растений 2007, 144: 1763–1776. DOI: 10.1104 / pp.107.103762 DOI: 10.1104 / pp.107.103762 10.1104 / pp.107.103762
Артикул Google Scholar
Гао Л., Ши YX: Генетические различия в устойчивости арахиса к железодефицитному хлорозу. J Plant Nutr 2007, 30: 37–52. DOI: 10.1080 / 01
CAS Статья Google Scholar
Graziano M, Lamattina L: Оксид азота и железо в растениях: новая и развивающаяся история. Trends Plant Sci 2005, 10: 4–8. doi: 10.1016 / j.tplants.2004.12.004 doi: 10.1016 / j.tplants.2004.12.004 10.1016 / j.tplants.2004.12.004
CAS Статья PubMed Google Scholar
Graziano M, Lamattina L: Накопление оксида азота необходимо для молекулярных и физиологических реакций на дефицит железа в корнях томатов. Завод J 2007, 52: 949–960.doi: 10.1016 / S0925–8388 (97) 00457-X doi: 10.1016 / S0925-8388 (97) 00457-X 10.1111 / j.1365-313X.2007.03283.x
Завод J 2007, 52: 949–960.doi: 10.1016 / S0925–8388 (97) 00457-X doi: 10.1016 / S0925-8388 (97) 00457-X 10.1111 / j.1365-313X.2007.03283.x
CAS Статья PubMed Google Scholar
Graziano M, Beligni MV, Lamattina L: Оксид азота улучшает внутреннюю доступность железа в растениях. Физиология растений 2002, 130: 1852–1859. http://dx.doi.org/10.1104/pp.009076 10.1104 / pp.009076
CAS Статья PubMed PubMed Central Google Scholar
Gunes A, Inal A, Alpaslan M, Eraslan F, Bagci EG, Cicek N: Салициловая кислота вызвала изменения некоторых физиологических параметров, симптоматичных для окислительного стресса и минерального питания кукурузы (Zea mays L.) выращенные при засолении. J Plant Physiol 2007, 164: 728–736. DOI: 10.1016 / j.jplph.2005.12.009 DOI: 10.1016 / j.jplph.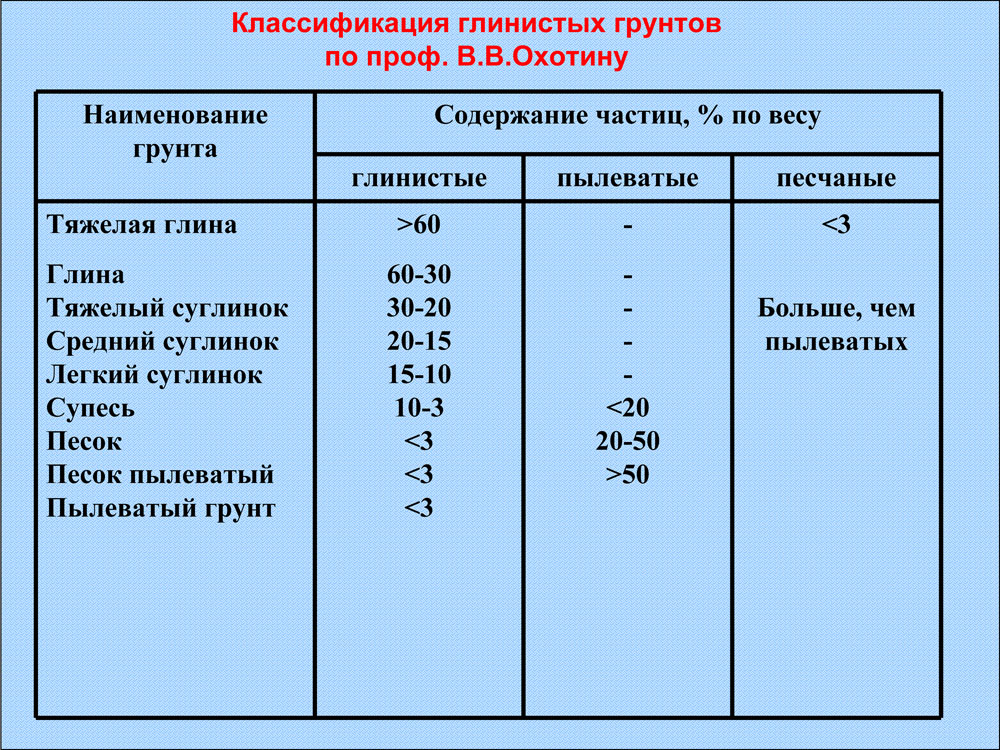 2005.12.009 10.1016 / j.jplph.2005.12.009
2005.12.009 10.1016 / j.jplph.2005.12.009
CAS Статья PubMed Google Scholar
Guo B, Liang YC, Zhu YG, Zhao FJ: Роль салициловой кислоты в облегчении окислительного повреждения корней риса (Oryza sativa), подвергнутых кадмиевому стрессу. Загрязнение окружающей среды 2007, 147: 743–749.doi: 10.1016 / j.envpol.2006.09.007 doi: 10.1016 / j.envpol.2006.09.007 10.1016 / j.envpol.2006.09.007
CAS Статья PubMed Google Scholar
Hager A: Роль плазматической мембраны H + -АТФаза в ауксин-индуцированном элонгационном росте: исторические и новые аспекты. Journal of Plant Res 2003, 116: 483–505. DOI: 10.1007 / s10265–003–0110-x doi: 10.1007 / s10265-003-0110-x 10.1007 / s10265-003-0110-x
CAS Статья Google Scholar
Hakan CA, Vahap K: Некоторые параметры, касающиеся статуса питания железом персиковых садов. J Biol Environ Sci 2007, 1: 111–115.
J Biol Environ Sci 2007, 1: 111–115.
Google Scholar
He JY, Ren YF, Pan XB, Yan YP, Zhu C, Jiang D: Салициловая кислота снижает токсическое действие кадмия на прорастание, рост проростков и активность амилазы риса. J Plant Nutr Soil Sci 2010, 173: 300–305. DOI: 10.1002 / jpln.200800302 doi: 10.1002 / jpln.200800302 10.1002 / jpln.200800302
CAS Статья Google Scholar
Heath RL, Packer L: Фотопероксидация в изолированных хлоропластах. I. Кинетика и стехиометрия перекисного окисления жирных кислот. Arch Biochem Biophys 1968, 125: 189–198. DOI: 10.1016 / 0003–9861 (68) –1 DOI: 10.1016 / 0003-9861 (68) -1 10.1016 / 0003-9861 (68) -1
CAS Статья PubMed Google Scholar
Hoagland DR, Arnon DI: Метод водного культивирования для выращивания растений без почвы. Круговой. Калифорния AES 1950, 347: 29–32.
Круговой. Калифорния AES 1950, 347: 29–32.
Google Scholar
Хорват Э., Салаи С., Джанда Т: Индукция устойчивости к абиотическому стрессу с помощью передачи сигналов салициловой кислоты. J Регулятор роста растений 2007, 26: 290–300. DOI: 10.1007 / s00344–007–9017–4 DOI: 10.1007 / s00344-007-9017-4 10.1007 / s00344-007-9017-4
CAS Статья Google Scholar
Hu KD, Hu LY, Li YH, Zhang FQ, Zhang H: Защитные роли оксида азота в прорастании и антиоксидантном метаболизме семян пшеницы в условиях медного стресса. Регулятор роста растений 2007, 53: 173–183.DOI: 10.1007 / s10725–007–9216–9 DOI: 10.1007 / s10725-007-9216-9 10.1007 / s10725-007-9216-9
CAS Статья Google Scholar
Jin CW, He XY, Wu P, Zheng SJ: Механизмы микробиологического накопления Fe в клевере луговом (Trifolium pretense L. ). Среда растительных клеток 2006, 29: 888–897. DOI: 10.1111 / j.1365–3040.2005.01468.x DOI: 10.1111 / j.1365-3040.2005.01468.x 10.1111 / j.1365-3040.2005.01468.x
). Среда растительных клеток 2006, 29: 888–897. DOI: 10.1111 / j.1365–3040.2005.01468.x DOI: 10.1111 / j.1365-3040.2005.01468.x 10.1111 / j.1365-3040.2005.01468.x
Артикул PubMed Google Scholar
Jin CW, You GY, He YF, Tang CX, Wu P, Zheng SJ: Вызванная дефицитом железа секреция фенольных соединений способствует повторному использованию апопластного железа корней клевера красного. Физиология растений 2007, 144: 278–285. DOI: 10.1104 / pp.107.095794 DOI: 10.1104 / pp.107.095794 10.1104 / pp.107.095794
CAS Статья PubMed PubMed Central Google Scholar
Карлидаг Х., Йилдирим Э., Туран М: Экзогенное применение салициловой кислоты влияет на качество и урожайность клубники, выращенной в теплицах с подогревом против замораживания. J Plant Nutr Soil Sci 2009, 172: 270–276. doi: 10.1002 / jpln.200800058 doi: 10.1002 / jpln.200800058 10.1002 / jpln.200800058
doi: 10.1002 / jpln.200800058 doi: 10.1002 / jpln.200800058 10.1002 / jpln.200800058
CAS Статья Google Scholar
Каземи Н., Хавари-Неджад Р.А., Фахими Х., Саадатманд С., Неджад-Саттари T: Влияние экзогенной салициловой кислоты и оксида азота на перекисное окисление липидов и активность антиоксидантных ферментов в листьях Brassica napus L. при никелевом стрессе. Sci Hortic-Amsterdam 2010, 126: 402–407. DOI: 10.1016 / j.scienta2010.07.037 DOI: 10.1016 / j.scienta2010.07.037 10.1016 / j.scienta.2010.07.037
CAS Статья Google Scholar
Хан В., Притхивирадж Б., Смит DL: Фотосинтетические реакции кукурузы и сои на некорневую подкормку салицилатов. J Plant Physiol 2003, 160: 485–492. DOI: 10.1078 / 0176–1617–00865 DOI: 10.1078 / 0176-1617-00865 10.1078 / 0176-1617-00865
CAS Статья PubMed Google Scholar
Ходары SEA: Влияние салициловой кислоты на рост, фотосинтез и углеводный обмен у растений кукурузы, подвергшихся солевому стрессу. Int J Agric Biol 2004, 6: 5–8.
Int J Agric Biol 2004, 6: 5–8.
CAS Google Scholar
Klessig DF, Durner J, Noad R, Navarre DA, Wendehenne D, Kumar D, Zhou JM, Shali S, Zhang S, Kachroo P, Trifa Y, Pontier D, Lam E, Silva H: оксид азота и передача сигналов салициловой кислоты при защите растений. Proc Natl Acad Sci U S A 2000, 97: 8849–8855. DOI: 10.1073 / pnas.97.16.8849 DOI: 10.1073 / pnas.97.16.8849 10.1073 / pnas.97.16.8849
CAS Статья PubMed PubMed Central Google Scholar
Knudson LL, Tibbitts TW, Edwards GE: Измерение озонового повреждения путем определения концентрации хлорофилла в листьях. Физиология растений 1977, 60: 606–608.doi: 10.1104 / pp.60.4.606 doi: 10.1104 / pp.60.4.606 10.1104 / pp.60.4.606
CAS Статья PubMed PubMed Central Google Scholar
Лигаба А. , Ямагути М., Шен Х, Сасаки Т., Ямамото Ю., Мацумото Н: Дефицит фосфора усиливает активность Н + -АТФазы плазматической мембраны и экссудацию цитрата в большом пурпурном люпине (Lupinus pilosus). Funct Plant Biol 2004, 31: 1075–1083. DOI: 10.1071 / FP04091 DOI: 10.1071 / FP04091 10.1071 / FP04091
, Ямагути М., Шен Х, Сасаки Т., Ямамото Ю., Мацумото Н: Дефицит фосфора усиливает активность Н + -АТФазы плазматической мембраны и экссудацию цитрата в большом пурпурном люпине (Lupinus pilosus). Funct Plant Biol 2004, 31: 1075–1083. DOI: 10.1071 / FP04091 DOI: 10.1071 / FP04091 10.1071 / FP04091
CAS Статья Google Scholar
Максвелл К., Джонсон GN: Флуоресценция хлорофилла — практическое руководство. J Exp Bot 2000, 51: 659–668. doi: 10.1093 / jexbot / 51.345.659 doi: 10.1093 / jexbot / 51.345.659 10.1093 / jexbot / 51.345.659
CAS Статья PubMed Google Scholar
Metwally A, Finkemeier I, George M, Dietz K: Салициловая кислота снижает токсичность кадмия в семенах ячменя. Физиология растений 2003, 132: 272–281. DOI: 10.1104 / pp.102.018457 DOI: 10.1104 / pp.102.018457 10.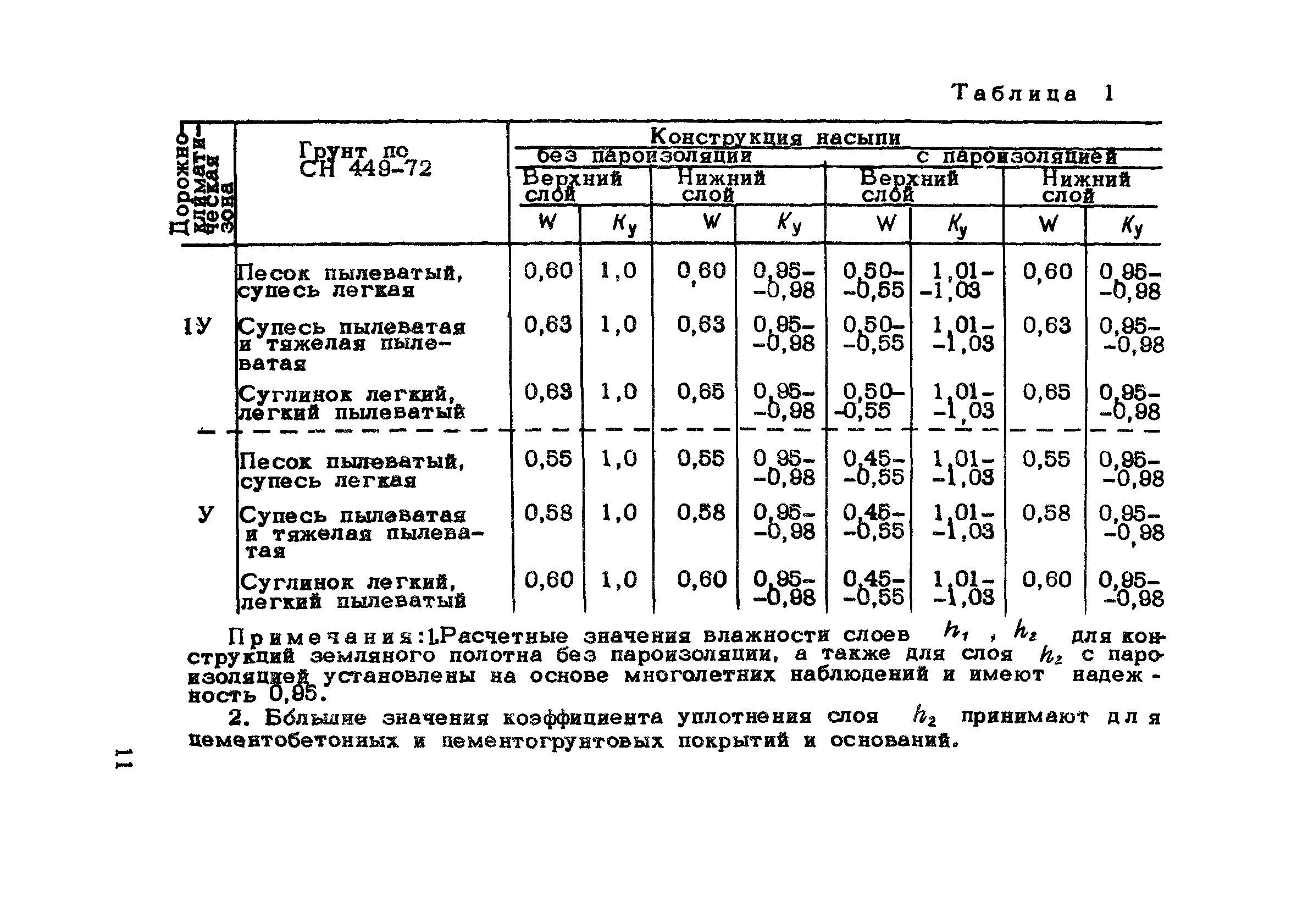 1104 / pp.102.018457
1104 / pp.102.018457
CAS Статья PubMed PubMed Central Google Scholar
Mittler R: Окислительный стресс, антиоксиданты и стрессоустойчивость. Trends Plant Sci 2002, 7: 405–410. DOI: 10,1016 / s1360–1385 (02) 02312–9 DOI: 10,1016 / s1360-1385 (02) 02312-9 10.1016 / S1360-1385 (02) 02312-9
CAS Статья PubMed Google Scholar
Molassiotis A, Therios I, Dimassi K, Diamantidis G, Chatzissavvidis C: Индукция активности Fe (III) -хелатредуктазы этиленом и салициловой кислотой в железодефицитных эксплантатах подвоя персика. J Plant Nutr 2005, 28: 669–682. DOI: 10.1081 / PLN-200052641 DOI: 10.1081 / PLN-200052641 10.1081 / PLN-200052641
CAS Статья Google Scholar
Morales F, Belkhodja R, Abadía A, Abadía J: Эффективность Фотосистемы II и механизмы рассеивания энергии в железодефицитных, выращиваемых в полевых условиях деревьях груши (Pyrus communis L.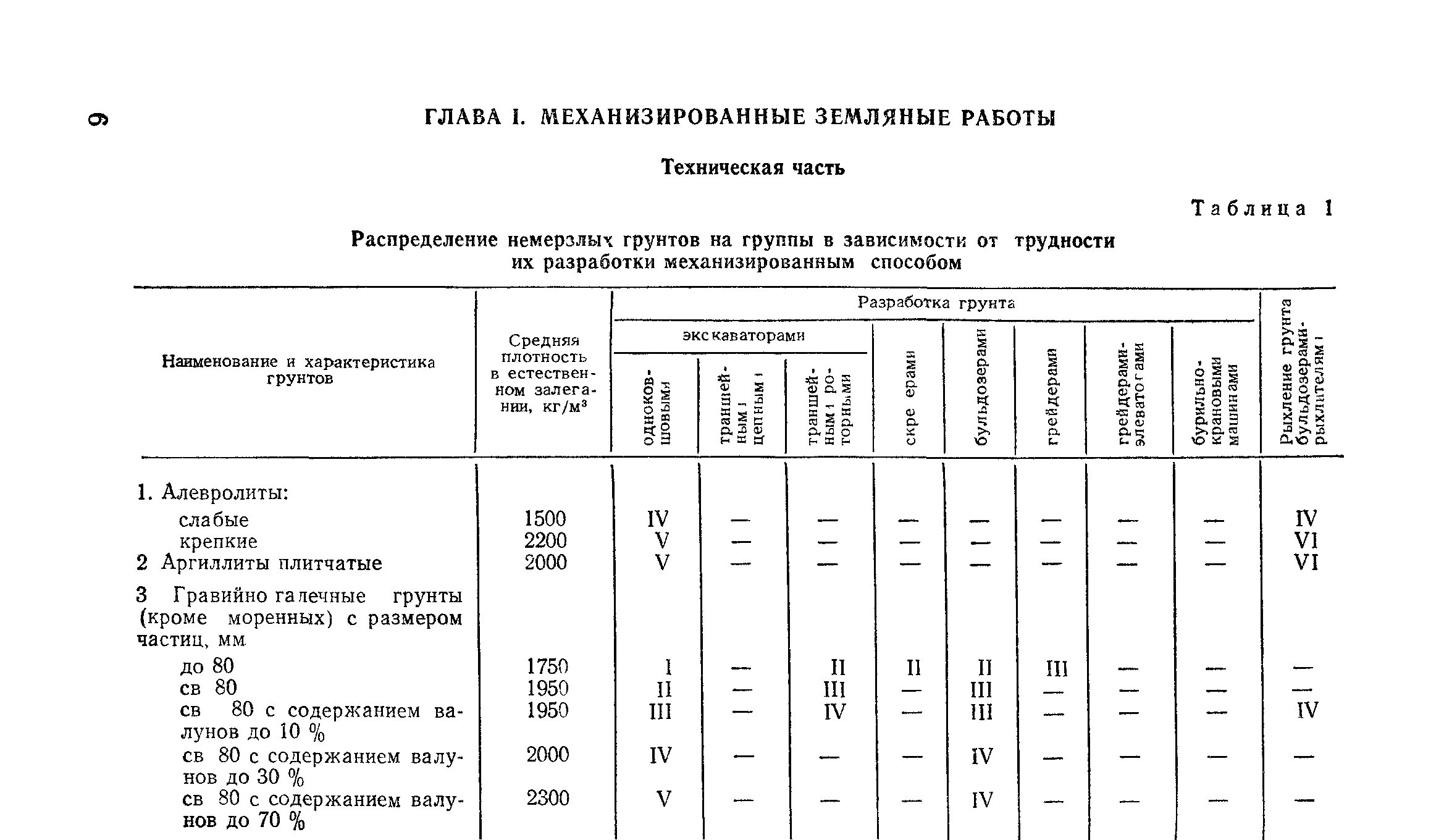 ). Photosynth Res 2000, 63: 9–21. DOI: 10.1023 / A.1006389
). Photosynth Res 2000, 63: 9–21. DOI: 10.1023 / A.1006389
CAS Статья PubMed Google Scholar
Никель К.С., Каннингем Б.А.: Улучшенный метод анализа пероксидазы с использованием лейко 2,3,6-трихлориндофенола и его применение для сравнительных измерений катализа пероксидазой. Ann Clin Biochem 1969, 27: 292–299.10.1016 / 0003-2697 (69) -9
CAS Статья Google Scholar
Охиниши Т., Галл Р.С., Майер М.Л.: Улучшенный анализ неорганического фосфата в присутствии экстралабильных фосфатных соединений: применение к анализу АТФ-азы в присутствии фосфокреатина. Anal Biochem 1975, 69: 261–267. 10.1016 / 0003-2697 (75)
Артикул Google Scholar
Osses LR, Godoy CA: Характеристика плазматической мембраны H + -АТФаза в двух разновидностях кофейных листьев (Coffea arabica L. ) и его взаимодействие с элиситорной фракцией гриба апельсиновой ржавчины (H. Vastatrix Berk and Br.) расы II. Plant Physiol Bioch 2006, 44: 226–235. doi: 10.1016 / j.plaphy.2006.01.009 doi: 10.1016 / j.plaphy.2006.01.009 10.1016 / j.plaphy.2006.01.009
) и его взаимодействие с элиситорной фракцией гриба апельсиновой ржавчины (H. Vastatrix Berk and Br.) расы II. Plant Physiol Bioch 2006, 44: 226–235. doi: 10.1016 / j.plaphy.2006.01.009 doi: 10.1016 / j.plaphy.2006.01.009 10.1016 / j.plaphy.2006.01.009
CAS Статья Google Scholar
Rao KVM, Sresty TVS: Антиоксидантные параметры проростков голубиного гороха (Cajanus cajan L. Millspaugh) в ответ на стрессы Zn и Ni. Plant Sci 2000, 157: 113–128. doi: 10.1016 / s0168–9452 (00) 00273–9 doi: 10.1016 / s0168-9452 (00) 00273-9 10.1016 / S0168-9452 (00) 00273-9
Статья Google Scholar
Раскин И .: Роль салициловой кислоты в растениях. Ann Rev Plant Physiol Mol Bio 1992, 43: 439–463. DOI: 10.1146 / annurev.pp.43.060192.002255 DOI: 10.1146 / annurev.pp.43.060192.002255 10.1146 / annurev. стр.43.060192.002255
стр.43.060192.002255
CAS Статья Google Scholar
Santi S, Schmidt W: Рассечение экструзии протонов, вызванной дефицитом железа, в корнях Arabidopsis. New Phytol 2009, 183: 1072–1084. DOI: 10.1111 / j.1469–8137.2009.02908.x DOI: 10.1111 / j.1469-8137.2009.02908.x 10.1111 / j.1469-8137.2009.02908.x
CAS Статья PubMed Google Scholar
Shi QH, Zhu ZJ: Влияние экзогенной салициловой кислоты на токсичность марганца, содержание элементов и антиоксидантную систему в огурце. Environ Exp Bot 2008, 63: 317–326. doi: 10.1016 / j.envexpbot.2007.11.003 doi: 10.1016 / j.envexpbot.2007.11.003 10.1016 / j.envexpbot.2007.11.003
CAS Статья Google Scholar
Shi GR, Cai QS, Liu QQ, Wu L: Снижение токсичности кадмия в растениях конопли, опосредованное салициловой кислотой, в отношении поглощения кадмия, фотосинтеза и антиоксидантных ферментов. Acta Physio Plant 2009, 31: 969–977.DOI: 10.1007 / s11738–009–0312–5 DOI: 10.1007 / s11738-009-0312-5 10.1007 / s11738-009-0312-5
Acta Physio Plant 2009, 31: 969–977.DOI: 10.1007 / s11738–009–0312–5 DOI: 10.1007 / s11738-009-0312-5 10.1007 / s11738-009-0312-5
CAS Статья Google Scholar
Simaei M, Khavarinejad RA, Saadatmand S, Bernard F, Fahimi H: Взаимодействие салициловой кислоты и оксида азота на растения сои при засолении NaCl. Russ J Plant Physiol 2011, 58: 783–790. DOI: 10.1134 / s1021443711050220 стр. 783–790 doi: 10.1134 / s1021443711050220 стр.783-790 10.1134 / S1021443711050220
CAS Статья Google Scholar
Sun BT, Jing Y, Chen KM, Song LL, Chen FJ, Zhang LX: Защитный эффект оксида азота на вызванный дефицитом железа окислительный стресс у кукурузы (Zea mays). J Plant Physiol 2007, 164: 536–543. doi: 10.1016 / j.jplph.2006.02.011 doi: 10.1016 / j.jplph.2006.02.011 10.1016 / j.jplph.2006. 02.011
02.011
CAS Статья PubMed Google Scholar
Vert G, Grotz N, Dedaldechamp F, Gaymard F, Guerinot ML, Briat JF, Curie C: IRT1, переносчик арабидопсиса, необходимый для поглощения железа из почвы и роста растений. Растительная клетка 2002, 14: 1223–1233. doi: 10.1105 / tpc.001388 doi: 10.1105 / tpc.001388 10.1105 / tpc.001388
CAS Статья PubMed PubMed Central Google Scholar
Йилдирим Э., Туран М., Гувенц I: Влияние обработки листовой салициловой кислотой на рост, содержание хлорофилла и минералов в огурце (Cucumis sativus L.), выращенном в условиях солевого стресса. J Plant Nutr 2008, 31: 593–612.10.1080 / 01
CAS Статья Google Scholar
Zhang XW, Zhang M, Wang QH, Qiu XK, Hu GQ, Dong YJ: Влияние экзогенного оксида азота на физиологические характеристики арахиса в условиях дефицита железа. J Plant Nutr and Fert 2011, 17: 665–673. http://www.plantnutrifert.org/CN/abstract/abstract2705.shtml
J Plant Nutr and Fert 2011, 17: 665–673. http://www.plantnutrifert.org/CN/abstract/abstract2705.shtml
CAS Google Scholar
Zhang XW, Dong YJ, Qiu XK, Hu GQ, Wang YH, Wang QH: Экзогенный оксид азота смягчает железодефицитный хлороз у арахиса, произрастающего на известняковой почве. Plant Soil Environ 2012, 58: 111–120.
CAS Google Scholar
Zottini M, Costa A, Michele RD, Ruzzene M, Carimi F, Schiavo FL: Салициловая кислота активирует синтез оксида азота в Arabidopsis. J Exp Bot 2007, 58: 1397–1405. DOI: 10.1093 / jxb / erm001 doi: 10.1093 / jxb / erm001 10.1093 / jxb / erm001
CAS Статья PubMed Google Scholar
Zuo YM, Zhang FS, Li XL, Cao YP: Исследования по улучшению железного питания арахиса путем пересева с кукурузой на известковой почве.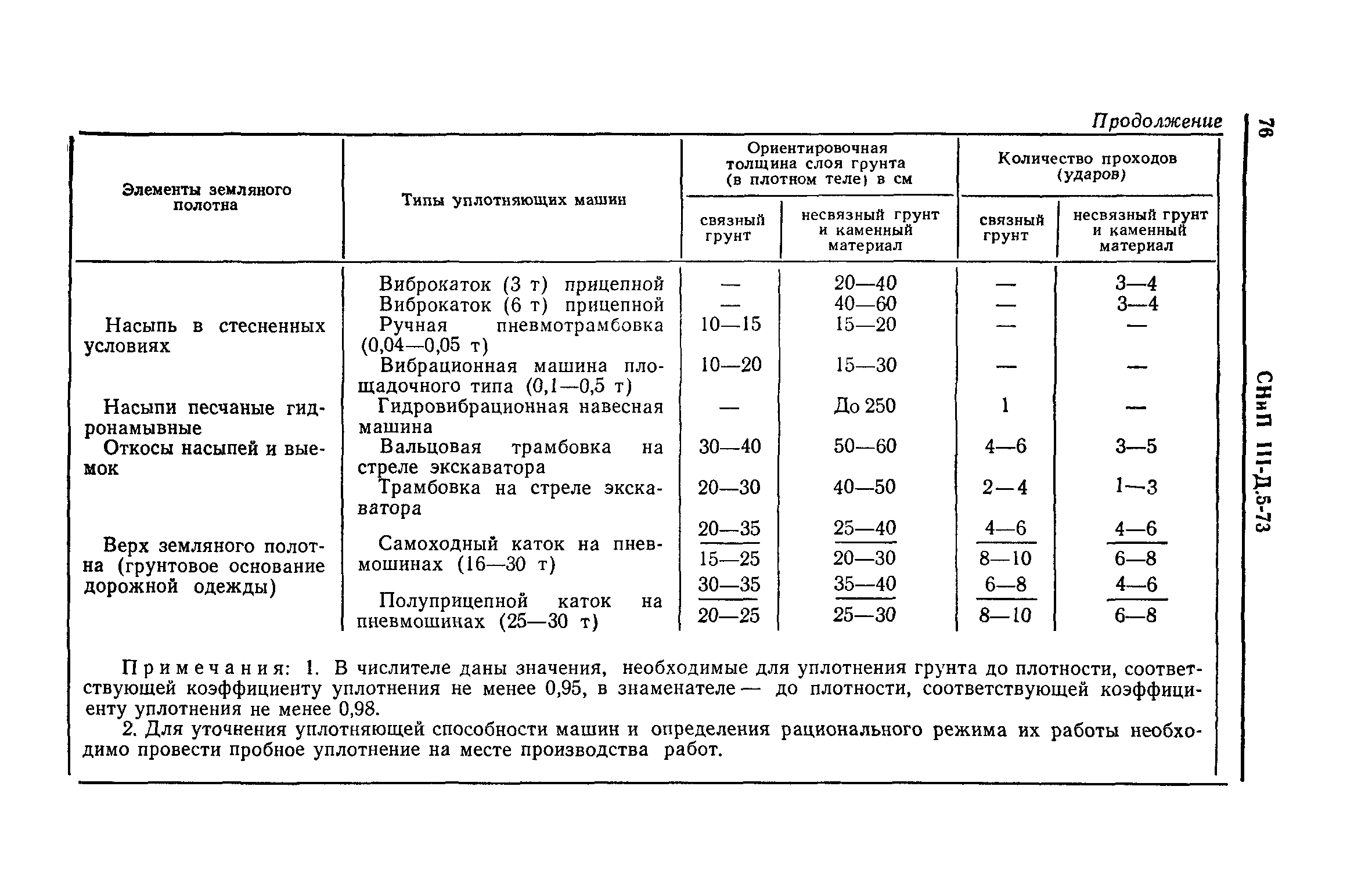 Почва растений 2000, 220: 13–25. DOI: 10.1023 / A: 1004724219988 DOI: 10.1023 / A: 1004724219988 10.1023 / A: 1004724219988
Почва растений 2000, 220: 13–25. DOI: 10.1023 / A: 1004724219988 DOI: 10.1023 / A: 1004724219988 10.1023 / A: 1004724219988
CAS Статья Google Scholar
Полногеномное ассоциативное картирование в разнообразной коллекции ярового ячменя выявляет наличие горячих точек QTL и генов-кандидатов для признаков архитектуры корня и побегов на стадии проростков | BMC Plant Biology
Abdel-Ghani AH. Селекция высокоурожайных линий разнородных староместных сортов иорданского ячменя в условиях сильного полива и стрессовых условий засухи.Вестник сельскохозяйственного факультета. Каирский университет Bull Fac Agric. 2013; (64): 13–29.
Солх М., ван Гинкель М. Готовность к засухе и смягчение ее последствий в засушливых районах развивающихся стран. Weather Clim Extrem. 2014; 3: 62–6.
Артикул Google Scholar
Продовольственная и сельскохозяйственная организация Объединенных Наций.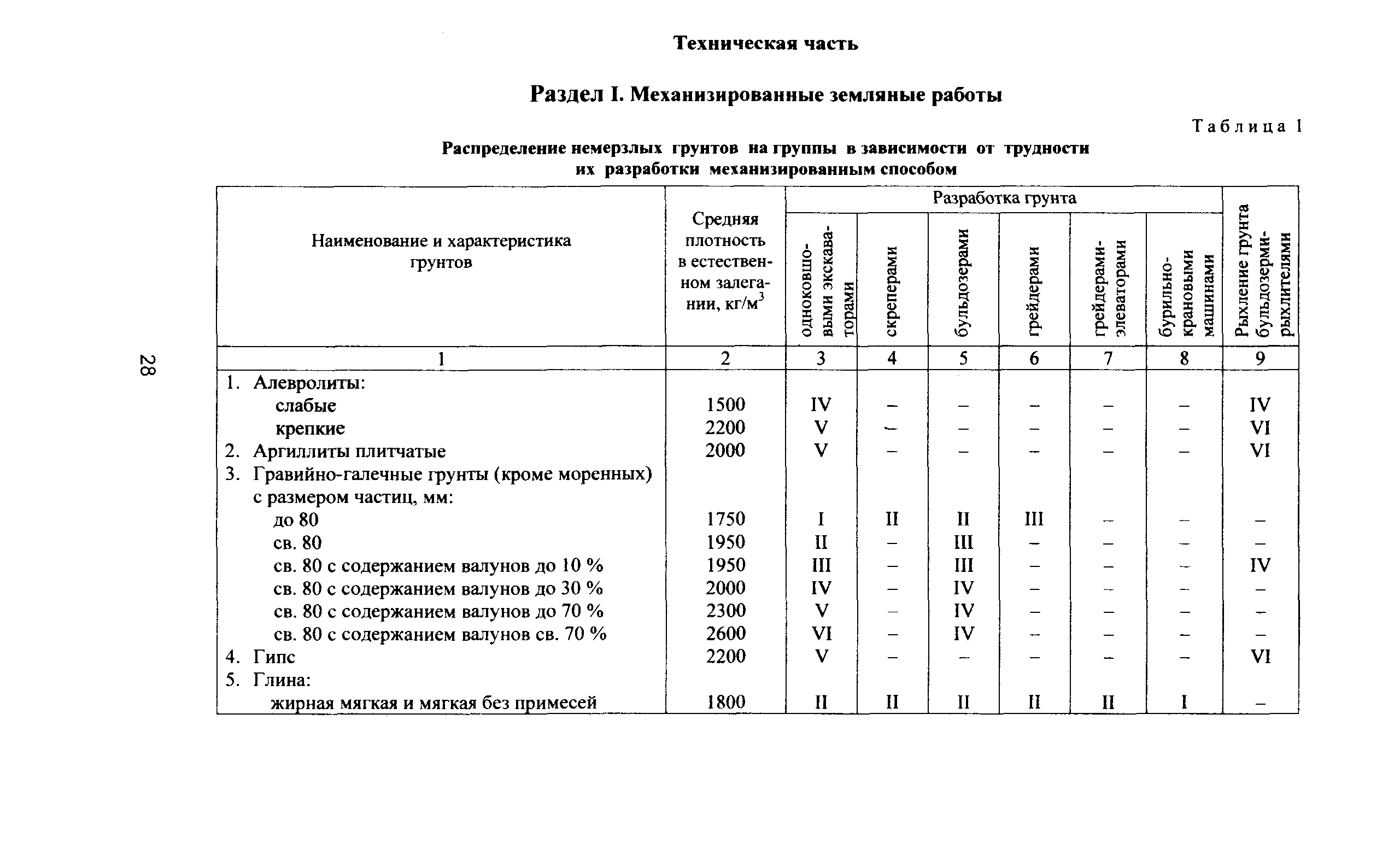 Статистический ежегодник ФАО за 2013 год.
Статистический ежегодник ФАО за 2013 год.
Чеккарелли С. Специфическая адаптация и разведение для маргинальных условий.Euphytica. 1994; 77: 205–19.
Артикул Google Scholar
Абдель-Гани А.Х., Кумар Б., Рейес-Матаморос Дж., Гонсалес-Портилья П.Дж., Янсен С., Мартин Дж. П.С. и др. Генотипическая изменчивость и взаимосвязь между признаками проростков и взрослых растений у инбредных линий кукурузы ( Zea mays L.), выращенных при контрастных уровнях азота. Euphytica. 2012. 189 (1): 123–33.
Артикул CAS Google Scholar
Шавруков Ю., Куришбаев А., Джатаев С., Швидченко В., Зотова Л., Кекемер Ф. и др. Раннее цветение как механизм защиты растений от засухи: как оно может помочь производству пшеницы? Фронтальный завод им. 2017; 8: 1950.
Артикул PubMed PubMed Central Google Scholar
Аль-Караки GN. Реакция пшеницы и ячменя во время прорастания на осмоподготовку семян при разном водном потенциале. J Agron Crop Sci. 1998. 181: 229–35.
Артикул Google Scholar
Хаддадин М.Ф., Абдель-Гани А.Х., Аль-Маджали Н.Ф. Реакция сортов на засуху на разных стадиях развития. Иордания Дж. Сельскохозяйственные науки. 2013; 9: 507–24.
Google Scholar
Richards RA, Passioura JB. Морфология семенных корней и водопользование пшеницы I. Воздействие на окружающую среду 1. Crop Sci. 1981; 21: 249.
Артикул Google Scholar
Grando S, Ceccarelli S.Морфология семенных корней и длина колеоптилей дикого ( Hordeum vulgare ssp. spontaneum ) и культурного ( Hordeum vulgate ssp. vulgare ) ячменя. Euphytica. 1995; 86: 73–80.
Артикул Google Scholar

Кашиваги Дж., Кришнамурти Л., Упадхьяя HD, Кришна Х., Чандра С., Вадез В. и др. Генетическая изменчивость корневых признаков, позволяющих избегать засухи, в коллекции мини-ядерной зародышевой плазмы нута ( Cicer arietinum L.). Euphytica. 2005; 146: 213–22.
Артикул Google Scholar
Комас Л.Х., Беккер С.Р., Круз ВМВ, Бирн П.Ф., Дириг Д.А. Признаки корней, способствующие продуктивности растений в условиях засухи. Фронтальный завод им. 2013; 4: 442.
Артикул PubMed PubMed Central Google Scholar
Хеллал Ф.А., Эль-Шабрави Х.М., Абд Эль-Хади М., Хатаб И.А., Эль-Сайед САА, Абделли К.Влияние стресса засухи, вызванного ПЭГ, на молекулярные и биохимические составляющие и рост проростков египетских сортов ячменя. Журнал генной инженерии и биотехнологии. 2018; 16 (1): 203–12.
Артикул CAS PubMed Google Scholar
Хлоупек О., Достал В., Стршеда Т., Псота В., Дворжачкова О. Засухоустойчивость сортов ячменя в зависимости от размера их корневой системы. Порода растений. 2010. 129: 630–6.
Артикул Google Scholar
Sayed M. QTL-анализ засухоустойчивости, связанной с признаками корней и побегов ячменя ( Hordeum vulgare L.). Кандидатская диссертация Боннского университета. http://hss.ulb.uni-bonn.de/2011/2521/2521.pdf. 2011.
Чен Г., Кругман Т., Фахима Т., Чен К., Рёдер М., Нево Э., Корол А. Хромосомные области, контролирующие засухоустойчивость проростков израильского дикого ячменя, Hordeum spontaneum C. Koch. Genet Resour Crop Evol. 2010; 57: 85.
Артикул Google Scholar
Свачина П., Стршеда Т., Хлоупек О. Необычный отбор по размеру корневой системы увеличивает урожайность ячменя. Agron Sustain Dev. 2013; 34: 545–51.
Артикул Google Scholar
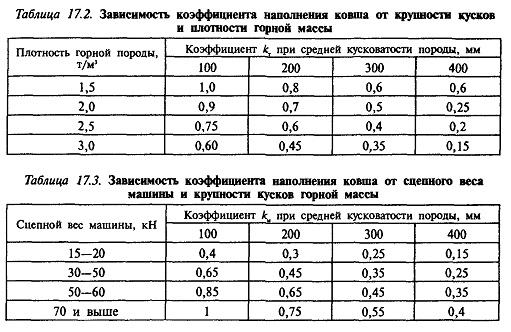
Аткинсон Дж. А., Винген Л. Ю., Гриффитс М., Паунд М. П., Гаджу О., Фоулкс М. Дж. И др. Фенотипирование показывает основной QTL роста корней проростков гексаплоидной пшеницы. J Exp Bot. 2015; 66 (8): 2283–92.
Артикул CAS PubMed PubMed Central Google Scholar
Maccaferri M, El-Feki W., Nazemi G, Salvi S, Canè MA, Colalongo MC и др. Приоритет локусов количественных признаков для архитектуры корневой системы тетраплоидной пшеницы. J Exp Bot. 2016; 67 (4): 1161–78.
Артикул CAS PubMed PubMed Central Google Scholar
Эндрю Р., Соланки С. Сравнительная морфология корней инбредных линий кукурузы в зависимости от производительности. Агрон Дж. 1966; 58: 415–8.
Артикул Google Scholar
Naz AA, Arifuzzaman M, Muzammil S, Pillen K, Léon J. Линии интрогрессии дикого ячменя выявили новые аллели QTL для корневых и связанных признаков побегов культурного ячменя ( Hordeum vulgare L. ). BMC Genet. 2014; 15: 107.
). BMC Genet. 2014; 15: 107.
Артикул PubMed PubMed Central Google Scholar
Сингх В., ван Остером Э., Джордан Д.Р., Мессина С.Д., Купер М., Хаммер Г.Л. Морфологическое и архитектурное развитие корневой системы сорго и кукурузы.Почва растений. 2010; 333: 287.
Артикул CAS Google Scholar
Zhu J, Kaeppler SM, Lynch JP. Картирование QTL для бокового ветвления и длины корня кукурузы ( Zea mays L.) при дифференциальной подаче фосфора. Theor Appl Genet. 2005; 111: 688–95.
Артикул CAS PubMed Google Scholar
Laperche A, Devienne-Barret F, Maury O, Le Gouis J, Ney B.Упрощенная концептуальная модель функционирования углерода / азота для анализа QTL адаптации озимой пшеницы к дефициту азота. Theor Appl Genet. 2006; 113: 1131–46.
Артикул CAS PubMed Google Scholar

Ли Х., Вайланкур Р., Мендхэм Н., Чжоу М. Сравнительное картирование локусов количественных признаков, связанных с толерантностью к заболачиванию ячменя ( Hordeum vulgare L.). BMC Genomics. 2008; 9: 401.
Артикул CAS PubMed PubMed Central Google Scholar
Лю X, Li R, Chang X, Jing R. Картирование QTL для признаков корня проростков в популяции удвоенной гаплоидной пшеницы при различных водных режимах. Euphytica. 2013; 189: 51–66.
Артикул Google Scholar
Хираман Д.А., Хопманс Дж. В., Клаусницер В. Трехмерная визуализация корней растений in situ с помощью рентгеновской компьютерной томографии. Почва растений. 1997; 189: 167–79.
CAS Google Scholar
Трейси С., Робертс Дж., Блэк С., Макнил А., Дэвидсон Р., Муни С. Х-фактор: визуализация ненарушенной корневой архитектуры в почвах с помощью рентгеновской компьютерной томографии. J Exp Bot. 2010; 61: 311–3.
J Exp Bot. 2010; 61: 311–3.
Артикул CAS PubMed Google Scholar
Муни С.Дж., Придмор Т.П., Хеллиуэлл Дж., Беннетт М.Дж. Разработка рентгеновской компьютерной томографии для неинвазивного визуализации трехмерной архитектуры корневой системы в почве. Почва растений. 2012; 352: 1–22.
Артикул CAS Google Scholar
Дейли К., Купер Л., Кеберник Н., Эваристо Дж., Киз С., ван Веелен А., Руз Т. Динамика воды в ризосфере. Ризосфера. 2017; 4: 139–51.
Артикул Google Scholar
Вальтер А., Шурр У. Динамика роста листьев и корней: эндогенный контроль в сравнении с воздействием окружающей среды. Энн Бот. 2005; 95: 891–900.
Артикул PubMed PubMed Central Google Scholar
Хоффманн А., Маурер А., Пиллен К. Обнаружение QTL дефицита азота в интрогрессионных линиях ювенильного дикого ячменя, растущих в гидропонной системе. BMC Genet. 2012; 13:88.
Обнаружение QTL дефицита азота в интрогрессионных линиях ювенильного дикого ячменя, растущих в гидропонной системе. BMC Genet. 2012; 13:88.
Артикул PubMed PubMed Central Google Scholar
Arifuzzaman M, Sayed MA, Muzammil S, Pillen K, Schumann H, Naz AA, Léon J. Обнаружение и проверка нового QTL для признаков побегов и корней ячменя ( Hordeum vulgare L.).Мол Порода. 2014; 34 (3): 1373–87.
Артикул CAS Google Scholar
Кумар Б., Абдель-Гани А.Х., Пейс Дж., Рейес-Матаморос Дж., Хоххолдингер Ф., Любберштедт Т. Ассоциативный анализ однонуклеотидных полиморфизмов в генах-кандидатах с корневыми признаками кукурузы ( Zea mays L.) саженцы. Plant Sci. 2014; 224: 9–19.
Артикул CAS PubMed PubMed Central Google Scholar
Tuberosa R, Salvi S, Sanguineti MC, Maccaferri M, Giuliani S, Landi P.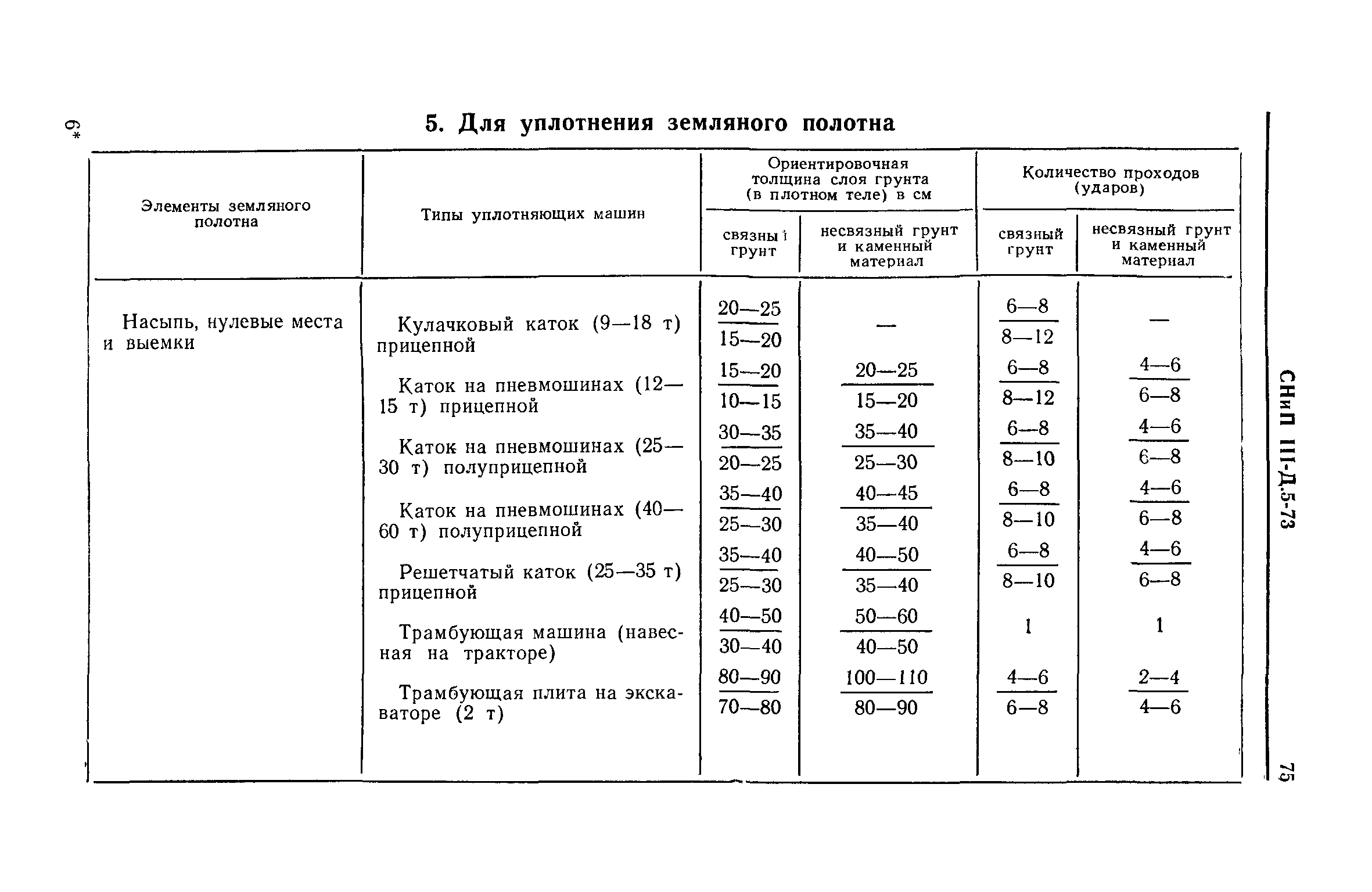 Поиск локусов количественных признаков, контролирующих корневые признаки кукурузы: критическая оценка. Почва растений. 2003. 255 (1): 35–54.
Поиск локусов количественных признаков, контролирующих корневые признаки кукурузы: критическая оценка. Почва растений. 2003. 255 (1): 35–54.
Артикул CAS Google Scholar
Baum M, Grando S, Backes G, Jahoor A, Sabbagh A, Ceccarelli S. QTL для агрономических признаков в средиземноморской среде, идентифицированных в рекомбинантных инбредных линиях кросса «Arta» x H.spontaneum 41-1. Theor Appl Genet. 2003; 107: 1215–25.
Артикул CAS PubMed PubMed Central Google Scholar
Ивандич В., Томас ВТБ, Нево Е., Чжан З., Форстер Б.П. Связь простых повторов последовательностей с вариациями количественных признаков, включая устойчивость к биотическим и абиотическим стрессам у Hordeum spontaneum. Порода растений. 2003. 122: 300–4.
Артикул CAS Google Scholar
Teulat B, Zoumarou-Wallis N, Rotter B, Ben Salem M, Bahri H, This D. QTL для относительного содержания воды в выращиваемом в поле ячменя и его стабильности в средиземноморской среде. Theor Appl Genet. 2003. 108 (1): 181–8.
QTL для относительного содержания воды в выращиваемом в поле ячменя и его стабильности в средиземноморской среде. Theor Appl Genet. 2003. 108 (1): 181–8.
Артикул CAS PubMed PubMed Central Google Scholar
Сюэ Д., Чжоу М., Чжан Х, Чен С., Вэй К., Цзэн Ф. и др. Определение QTL для урожайности и компонентов урожая ячменя в различных условиях роста.J Zhejiang Univ Sci B. 2010; 11 (3): 169–76.
Артикул CAS PubMed PubMed Central Google Scholar
Уолл К., Борсук Л.А., Странски Х., Нетлтон Д., Шнабл П.С., Хоххолдингер Ф. Изоляция, характеристика и анализ специфичных для перицикла транскриптомов нового мутанта инициации боковых и семенных корней кукурузы rum1 . Plant Physiol. 2005; 139: 1255–67.
Артикул CAS PubMed PubMed Central Google Scholar

Champoux MC, Wang G, Sarkarung S, Mackill DJ, O’Toole JC, Huang N, McCouch SR. Выявление генов, связанных с морфологией корней и предотвращением засухи у риса, посредством связывания с молекулярными маркерами. Theor Appl Genet. 1995; 90: 969–81.
Артикул CAS PubMed PubMed Central Google Scholar
Hetz W, Hochholdinger F, Schwall M, Feix G. Выделение и характеристика rtcs, мутанта кукурузы, у которого отсутствует образование узловых корней.Плант Дж. 1996; 10: 845–57.
Артикул CAS Google Scholar
Ван М., Цзян Н., Цзя Т., Лич Л., Кокрам Дж., Во Р. и др. Полногеномное ассоциативное картирование агрономических и морфологических признаков в высокоструктурированных популяциях сортов ячменя. Theor Appl Genet. 2012. 124 (2): 233–46.
Артикул PubMed PubMed Central Google Scholar
Neumann K, Kobiljski B, Denčić S, Varshney RK, Börner A.Полногеномное ассоциативное картирование: тематическое исследование мягкой пшеницы ( Triticum aestivum L.). Мол Порода. 2011; 27 (1): 37–58.
Артикул Google Scholar
Стич Б., Мелчингер А. Введение в ассоциативное картирование растений. CAB Rev.2010; 5: 1–9.
Артикул Google Scholar
Pasam RK, R. Sharma R, Malosetti M, van Eeuwijk FA, Haseneyer G, Kilian B, Graner A.Полногеномные ассоциативные исследования агрономических признаков во всемирной коллекции ярового ячменя. БМК Завод Биол 2012; 12:16.
Куртуа Б., Ахмади Н., Ховаджа Ф., Прайс А. Х., Рами Дж. Ф., Фруин Дж., Хамелин С., Руис М. Генетическая архитектура корня Райса: метаанализ из базы данных QTL засухи. Рис. 2009; 2: 115–28.
Артикул Google Scholar
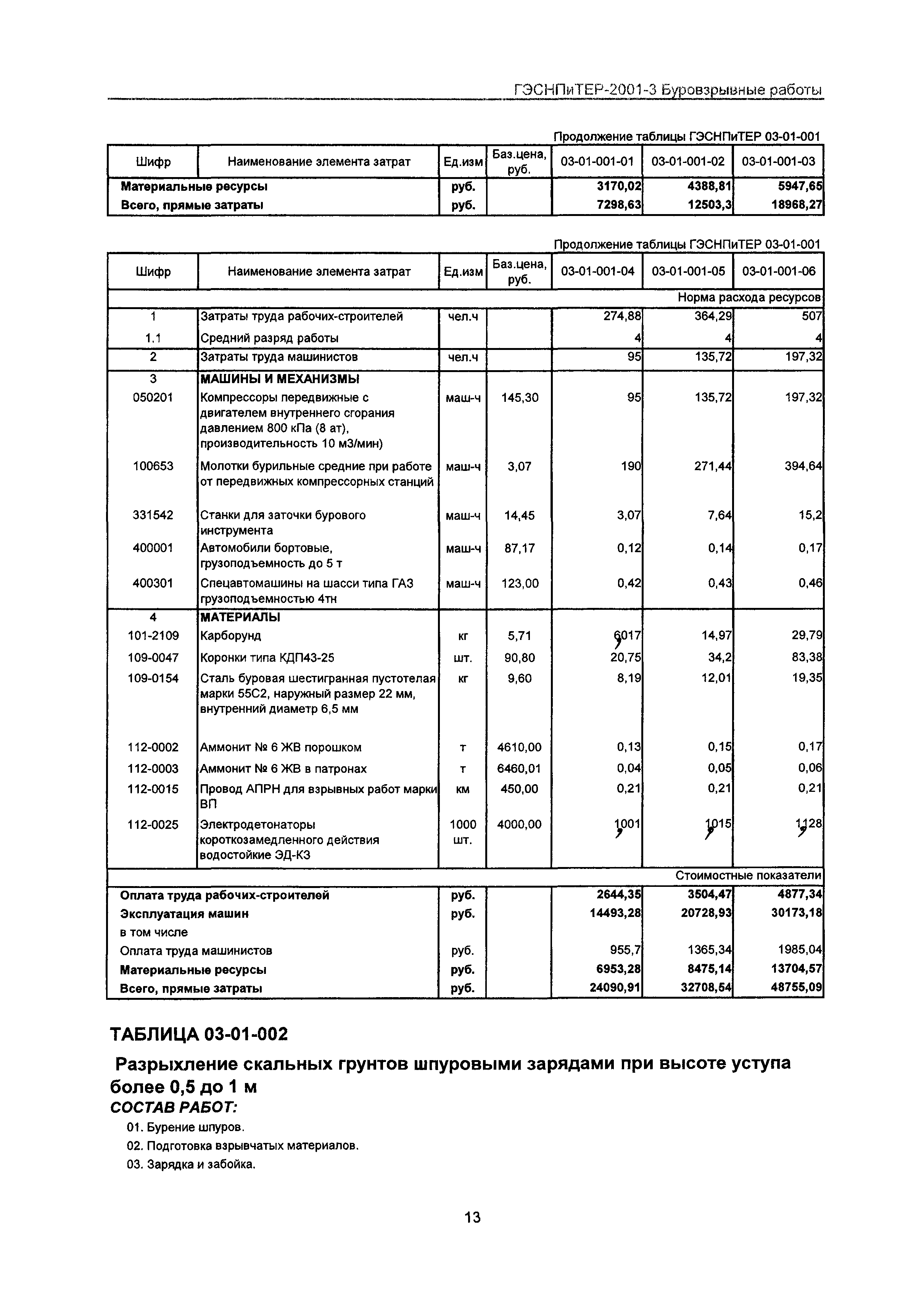
Ahmad Naz A, Ehl A, Pillen K, Léon J. Подтверждение корневых эффектов локуса количественных признаков дикого происхождения в культивируемом фоне ячменя ( Hordeum vulgare L.). Порода растений. 2012. 131 (3): 392–8.
Артикул Google Scholar
Зааре М., Джафари Х. Количественное разнообразие локусов признаков солеустойчивости на ранней стадии роста ячменя. Crop Breed J. 2013; 3: 69–77.
Google Scholar
Джордж Т.С., Браун Л.К., Рамзи Л., Уайт П.Дж., Ньютон А.С., Бенгоу А.Г., Рассел Дж., Томас В.Т.Б. Понимание генетического контроля и физиологических особенностей, связанных с производством ризошей ячменем ( Hordeum vulgare ).Новый Фитол. 2014; 203: 195–205.
Артикул CAS PubMed PubMed Central Google Scholar
Райнерт С., Корц А., Леон Дж., Наз А.А. Полногеномное ассоциативное картирование в наборе глобального разнообразия выявляет новый QTL, контролирующий корневую систему и связанные с ней вариации побегов ячменя. Фронтальный завод им. 2016; 19, 7.
Фронтальный завод им. 2016; 19, 7.
Шарма С., Сюй С., Эдаи Б., Хупс А., Клоуз Т.Дж., Лукашевский А.Дж., Уэйнес Дж. Рассмотрение эффектов QTL для корневых признаков с использованием популяции, специфичной для картирования хромосомных плеч мягкой пшеницы.Theor Appl Genet. 2011; 122: 759–69.
Артикул PubMed PubMed Central Google Scholar
Канэ М.А., Маккаферри М., Наземи Г., Салви С., Франсия Р., Колалонго С., Тубероза Р. Сопоставление архитектурных черт корня проростков твердой пшеницы с точки зрения агрономической эффективности. Мол Порода. 2014. 34 (4): 1629–45.
Артикул PubMed PubMed Central Google Scholar
Хамада А., Нитта М., Насуда С., Като К., Фудзита М., Мацунака и др. Новые QTL для угла роста семенных корней пшеницы ( Triticum aestivum L.). Почва растений 2012; 354: 395–405.

Цай Х., Чен Ф., Ми Г, Чжан Ф., Маурер Х.П., Лю В. и др. Картирование QTL для архитектуры корневой системы кукурузы ( Zea mays L.) в поле на разных стадиях развития. Theor Appl Genet. 2012; 125: 1313–24.
Артикул PubMed PubMed Central Google Scholar
Бертон А.Л., Джонсон Дж. М., Фёрстер Дж. М., Хирш К. Н., Буэлл С. Р., Кэпплер С. М., Браун К. М., Линч Дж. Картирование QTL и фенотипические вариации архитектурных признаков корня кукурузы ( Zea mays L.). Theor Appl Genet. 2014; 127: 2293–311.
Артикул PubMed PubMed Central Google Scholar
Хунд А., Реймер Р., Мессмер Р. Консенсусная карта QTL, контролирующих длину корня кукурузы. Почва растений. 2011; 344: 143–58.
Артикул CAS Google Scholar
Shelden MC, Roessner U, Sharp RE, Tester M, Bacic A. Генетическая изменчивость реакции роста корней генотипов ячменя на стресс засоления. Funct Plant Biol. 2013; 40 (5): 516–30.
Генетическая изменчивость реакции роста корней генотипов ячменя на стресс засоления. Funct Plant Biol. 2013; 40 (5): 516–30.
Артикул CAS Google Scholar
Long NV, Dolstra O, Malosetti M, Kilian B, Graner A, Visser RG, van der Linden CG. Ассоциативное картирование солеустойчивости ячменя ( Hordeum vulgare L.). Theor Appl Genet. 2013; 126: 2335–51.
Артикул CAS PubMed PubMed Central Google Scholar
Li SB, Xie ZZ, Hu CG, Zhang JT. Обзор факторов ответа на ауксин (ARF) у растений. Фронтальный завод им. 2016; 7: 47.
PubMed PubMed Central Google Scholar
Б. Ороса-Пуэнте Б., Лефтли Н., фон Вагенхайм Д., Банда Дж., Шривастава А.К., Хилл К. и др. Ветвление корней в сторону воды включает посттрансляционную модификацию фактора транскрипции ARF7.Наука. 2018; 362 (6421): 1407–1410.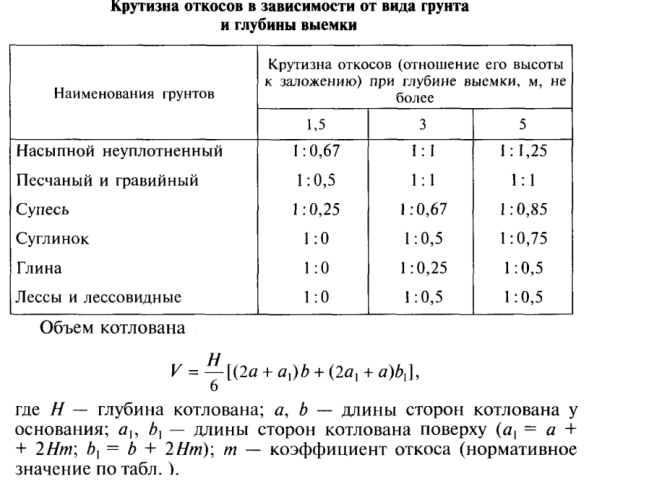
Tombuloglu H. Полногеномный анализ семейства генов факторов ответа на ауксин (ARF) ячменя ( Hordeum vulgare L.). J Plant Biochem Biotechnol. 2018; doi.org/10.1007/s13562-018-0458-6.
Uga Y, Sugimoto K, Ogawa S, Rane J, Ishitani M, Hara N, et al. Контроль архитектуры корневой системы с помощью DEEPER ROOTING 1 увеличивает урожай риса в условиях засухи. Нат Жене. 2013. 45 (9): 1097–102.
Артикул CAS PubMed PubMed Central Google Scholar
Квасьневский М., Зарейко И. Молекулярное клонирование и характеристика гена бета-экспансина, связанного с образованием корневых волосков у ячменя. Plant Physiol. 2006; 141: 1149–58.
Артикул CAS PubMed PubMed Central Google Scholar
Каур Р., Синг К., Синг Дж. Ген киназы, специфичной для корня, HvWAK1 , регулирует рост корня и сильно отличается у ячменя и других злаков. J Funct Integr Genomics.13 (2): 167–77.
J Funct Integr Genomics.13 (2): 167–77.
Войцеховски Т., Гудинг М.Дж., Рамзи Л., Грегори П.Дж. Влияние генов карликовости на рост корней проростков пшеницы. J Exp Bot. 2009. 60 (9): 2565–73.
Артикул CAS PubMed PubMed Central Google Scholar
Chloupek O, Forster BP, Thomas WTB. Влияние полукарликовых генов на размер корневой системы ячменя, выращиваемого в полевых условиях. Theor Appl Genet. 2006. 122 (5): 779–86.
Артикул CAS Google Scholar
Фосс-Фелс К.П., Робинсон Х., Мадж С.Р., Ричард С., Ньюман С., Витткоп Б. и др. ВЕРНАЛИЗАЦИЯ1 модулирует архитектуру корневой системы пшеницы и ячменя. Завод Мол. 2018; 11: 226–9.
Артикул CAS PubMed PubMed Central Google Scholar
Abdel-Ghani AH, Neumann K, Wabila C, Sharma R, Dhanagond S, Owais SJ, Börner A, Graner A, Kilian B.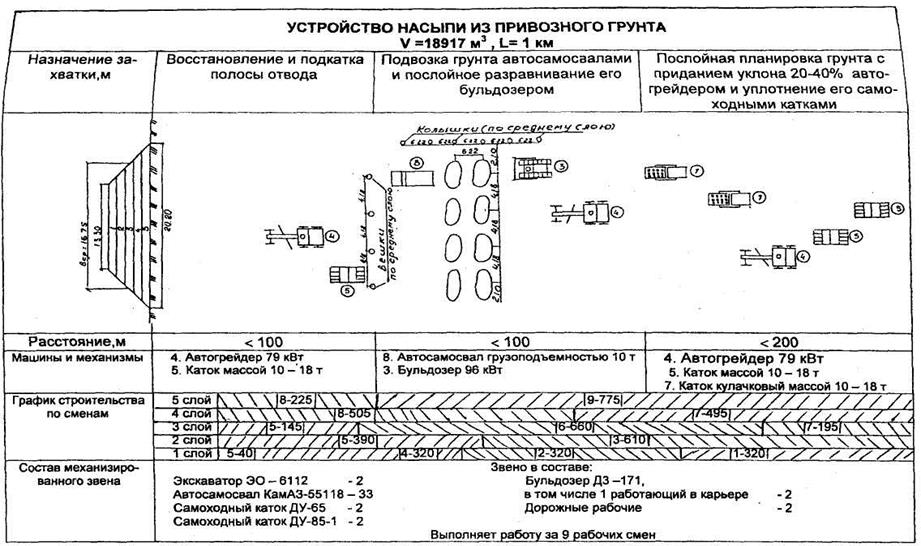 Л.) сбор в условиях моделирования засухи. Genet Resour Crop Evol. 2015; 62 (2): 275–92.
Л.) сбор в условиях моделирования засухи. Genet Resour Crop Evol. 2015; 62 (2): 275–92.
Артикул CAS Google Scholar
Mascher M, Gundlach H, Himmelbach A, Beier S, Twardziok SO, Wicker T, et al. Конформация хромосомы фиксирует упорядоченную последовательность генома ячменя. Природа. 2017; 544 (7651): 427–33.
Артикул CAS PubMed PubMed Central Google Scholar
Stracke S, Haseneyer G, Veyrieras J-B, Geiger HH, Sauer S, Graner A, Piepho H-P. Картирование ассоциаций выявляет действие и взаимодействие генов при определении времени цветения ячменя. Theor Appl Genet. 2009. 118 (2): 259–73.
Артикул CAS PubMed PubMed Central Google Scholar
Alqudah AM, Sharma R, Pasam RK, Graner A, Kilian B, Schnurbusch T. Генетическое расчленение фотопериодной реакции на основе gwas длительности фазы перед цветением ярового ячменя.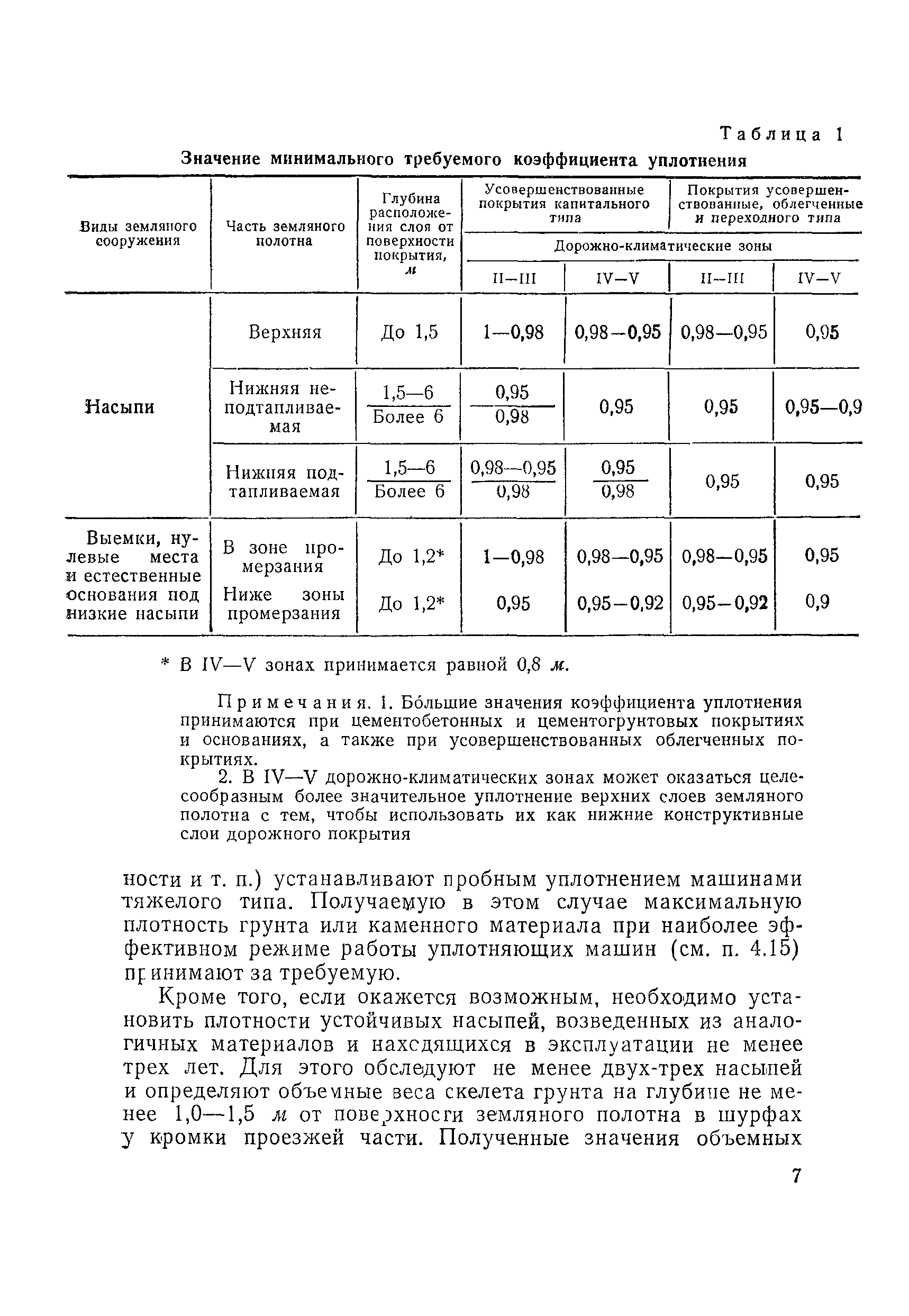 PLoS One. 2014; 9 (11): 1–27.
PLoS One. 2014; 9 (11): 1–27.
Артикул CAS Google Scholar
Yousseff HM, Eggert K, Koppolu R, Alqudah AM, Poursarebani N, Fazeli A, Sakuma S, Tagiri A, Rutten T., Govind G. VRS2 регулирует опосредованное гормонами формирование рисунка соцветий ячменя. Нат Жене. 2017; 49: 157–61.
Артикул CAS Google Scholar
Клукас С., Чен Д., Папе Дж. М.. Интегрированная аналитическая платформа: информационная система с открытым исходным кодом для высокопроизводительного фенотипирования растений.Plant Physiol. 2014; 165 (2): 506–18.
Артикул CAS PubMed PubMed Central Google Scholar
Hallauer AM, Miranda JB. Количественная генетика в селекции кукурузы. 2-е изд. Издательство государственного университета Айовы, Эймс, Айова, Соединенные Штаты Америки. 1981; 1–468.

R Основы статистических вычислений. R: язык и среда для статистических вычислений. R Основа статистических вычислений.Вена, Австрия; 2016. URL http://www.R-project.org/
Комадран Дж., Килиан Б., Рассел Дж., Рамзи Л., Стейн Н., Ганал М. и др. Естественные вариации гомолога Antirrhinum CENTRORADIALIS способствовали формированию весеннего роста и адаптации культурного ячменя к окружающей среде. Нат Жене. 2012; 44 (12): 1388–92.
Артикул CAS PubMed PubMed Central Google Scholar
Mascher M, Muehlbauer GJ, Rokhsar DS, Chapman J, Schmutz J, Barry K и др. Закрепление и упорядочение сборок контигов NGS путем популяционного секвенирования (POPSEQ). Плант Дж. 2013; 76: 718–27.
Артикул CAS PubMed PubMed Central Google Scholar
Тан И, Лю Х, Ван Дж, Ли М, Ван Кью, Тиан Ф и др. GAPIT версии 2: усовершенствованный интегрированный инструмент для геномной ассоциации и прогнозирования. Геном растений. 2016; 9 (2).
GAPIT версии 2: усовершенствованный интегрированный инструмент для геномной ассоциации и прогнозирования. Геном растений. 2016; 9 (2).
Wang Q, Tian F, Pan Y, Buckler ES, Zhang Z. Сверхмощный метод исследования ассоциации в целом по геному. PLoS One. 2014; 9 (9): e107684.
Артикул CAS PubMed PubMed Central Google Scholar
Бенджамини Ю., Хохберг Ю. Контроль уровня ложных открытий: практичный и эффективный подход к множественному тестированию. J R Stat Soc B. 1995; 57 (57): 289–300.
Google Scholar
Voorrips RE. MapChart: программное обеспечение для графического представления карт связей и QTL. J Hered. 2002. 93 (1): 77–8.
Артикул CAS PubMed Google Scholar
Colmsee C, Beier S, Himmelbach A, Schmutzer T., Scholz U, Mascher M. BARLEX — исследователь чернового генома ячменя. Завод Мол. 2015; 8: 964–6.
Завод Мол. 2015; 8: 964–6.
Артикул CAS PubMed Google Scholar
Милнер С.Г., Йост М., Такета С., Мазон Э.Р., Химмельбах А., Опперманн М. и др. Геномика генного банка подчеркивает разнообразие глобальной коллекции ячменя. Нат Жене. 2019; 51: 319–26.
Артикул CAS PubMed Google Scholar
Untergasser A, Cutcutache I, Koressaar T., Ye J, Faircloth BC, Remm M, Rozen SG. Primer3 — новые возможности и интерфейсы. Nucleic Acids Res. 2012; 40 (15): e115.
Артикул CAS PubMed PubMed Central Google Scholar
Холл ТА. BioEdit: удобный редактор выравнивания биологических последовательностей и программа анализа для Windows 95/98 / NT. Nucleic Acids Symp Ser. 1999; 41: 95–8.
CAS Google Scholar
Розас Дж. , Феррер-Мата А., Санчес-ДельБаррио Дж. К., Гирао-Рико С., Либрадо П., Рамос-Онсинс С.Е., Санчес-Грасиа А. DnaSP 6: анализ полиморфизма последовательностей ДНК для больших наборов данных. Mol Biol Evol. 2017; 34 (12): 3299–302.
, Феррер-Мата А., Санчес-ДельБаррио Дж. К., Гирао-Рико С., Либрадо П., Рамос-Онсинс С.Е., Санчес-Грасиа А. DnaSP 6: анализ полиморфизма последовательностей ДНК для больших наборов данных. Mol Biol Evol. 2017; 34 (12): 3299–302.
Артикул CAS PubMed Google Scholar
Neumann K, Zhao Y, Chu J, Keilwagen J, Reif JC, Kilian B, Graner A. Генетическая архитектура и временные закономерности накопления биомассы ярового ячменя, выявленные с помощью анализа изображений. BMC Plant Biol. 2017; 17 (1).
Венер Г., Балко С., Эндерс М., Хамбек К., Ордон Ф. Идентификация участков генома, участвующих в устойчивости к стрессу засухи и вызванному стрессом засухе старению листьев у ювенильного ячменя. BMC Plant Biol. 2015; 15: 125.
Артикул CAS PubMed PubMed Central Google Scholar
Мора Ф., Куитрал Ю.А., Матус И., Рассел Дж., Во Р.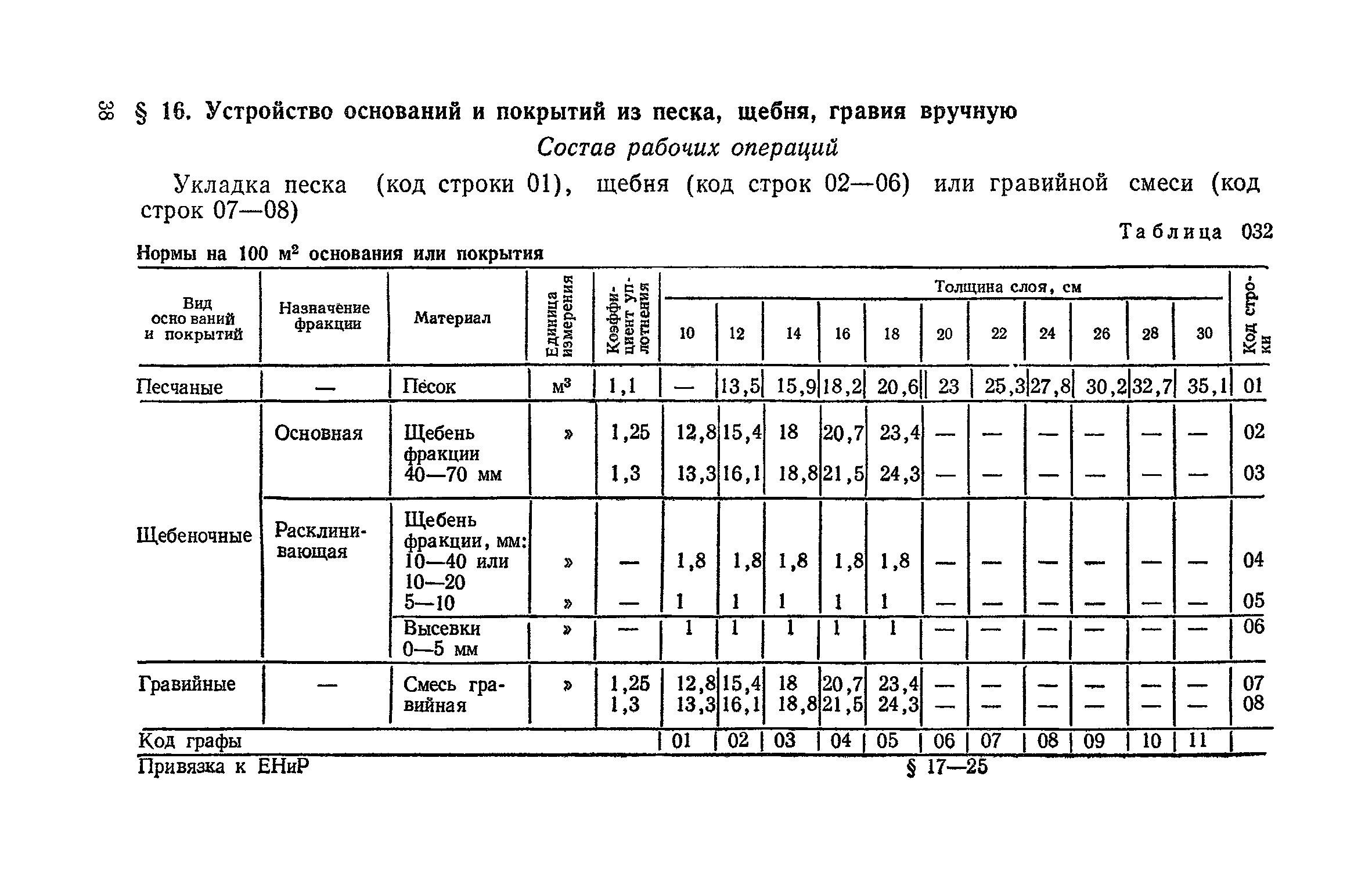 , дель Посо А. Картирование QTL на основе SNP 15 сложных признаков ячменя в неорошаемых и поливных условиях с помощью смешанного подхода к моделированию. Фронтальный завод им. 2016; 7: 909.
, дель Посо А. Картирование QTL на основе SNP 15 сложных признаков ячменя в неорошаемых и поливных условиях с помощью смешанного подхода к моделированию. Фронтальный завод им. 2016; 7: 909.
Артикул PubMed PubMed Central Google Scholar
Fletcher RS, Mullen JL, Heiliger A, McKay JK. QTL-анализ морфологии корней, времени цветения и урожайности выявляет компромиссы в ответ на засуху у Brassica napus .J Exp Bot. 2014; 66 (1): 245–56.
Артикул CAS PubMed PubMed Central Google Scholar
Дево Ю., Тоффано-Ниоче С., Клесс Дж., Теро В., Морин Н., Лауфс П. и др. Гены наиболее консервативной клады WOX у растений влияют на развитие корней и цветов арабидопсиса. BMC Evolutionary Biol. 2008; 8: 291.
Артикул CAS Google Scholar
Digel B, Tavakol E, Verderio G, Tondelli A, Xu X, Cattivelli L, Rossini L, von Korff M.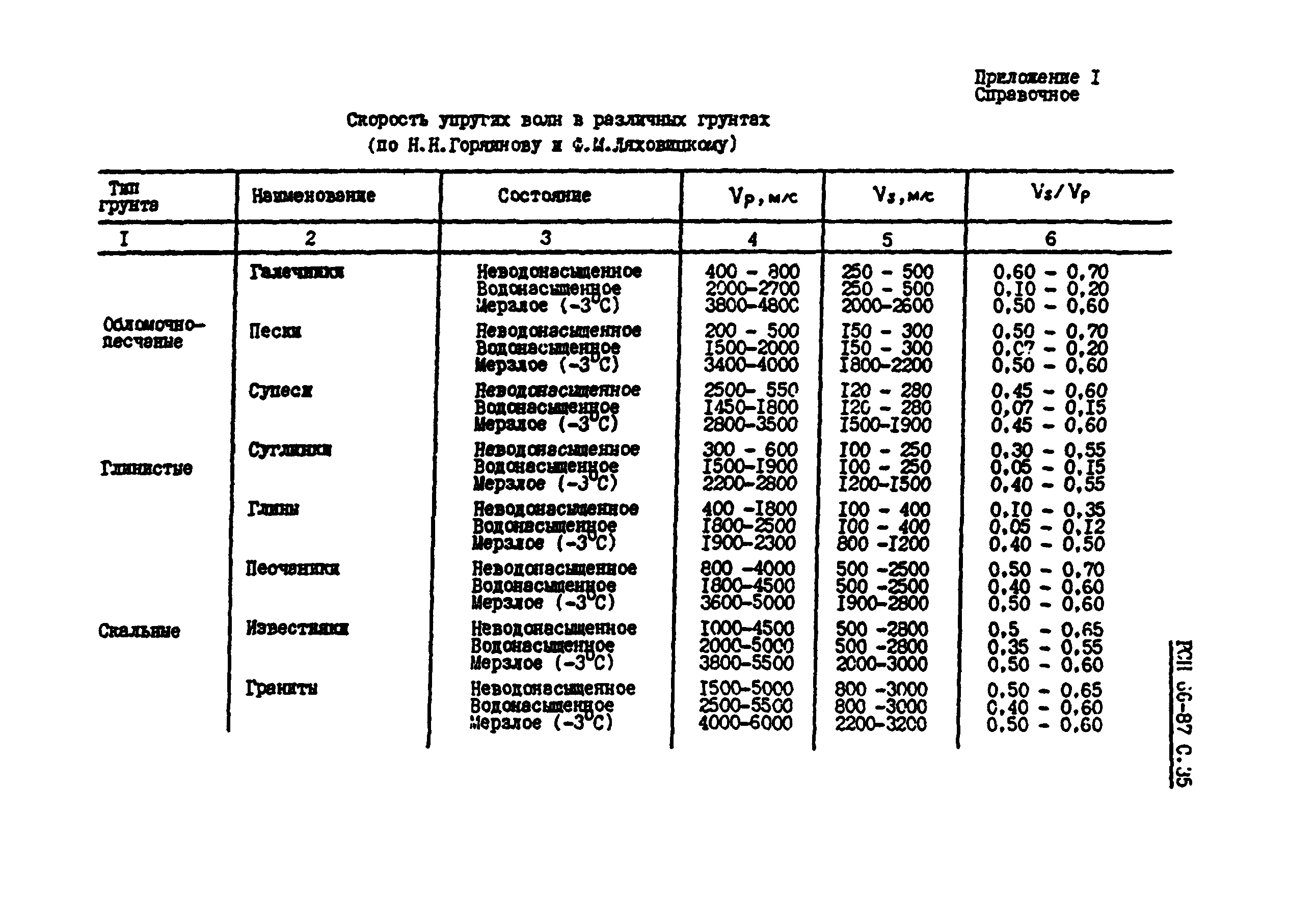 Photoperiod1 ( Ppd-h2 ) контролирует размер листьев. Plant Physiol. 2016; 172 (1): 405–15.
Photoperiod1 ( Ppd-h2 ) контролирует размер листьев. Plant Physiol. 2016; 172 (1): 405–15.
Артикул CAS PubMed PubMed Central Google Scholar
Habte E, Muller LM, Shtaya M, Davis SJ, Von Korff M. Осмотический стресс в корне ячменя влияет на экспрессию генов циркадных часов в побегах. Plant Cell Environ.2014; 37: 1321–7.
Артикул CAS PubMed Google Scholar
Мизуно Т., Ямашино Т. Сравнительный транскриптом генов с суточными колебаниями и гормонально-зависимых генов у Arabidopsis thaliana : понимание суточных реакций растений, контролируемых суточными часами, на обычные внешние стрессы. Physiol растительной клетки. 2008; 49: 481–7.
Артикул CAS PubMed Google Scholar
Müller A, Guan C, Gälweiler L, Tänzler P, Huijser P, Marchant A, Parry G, Bennett M, Wisman E, Palme K.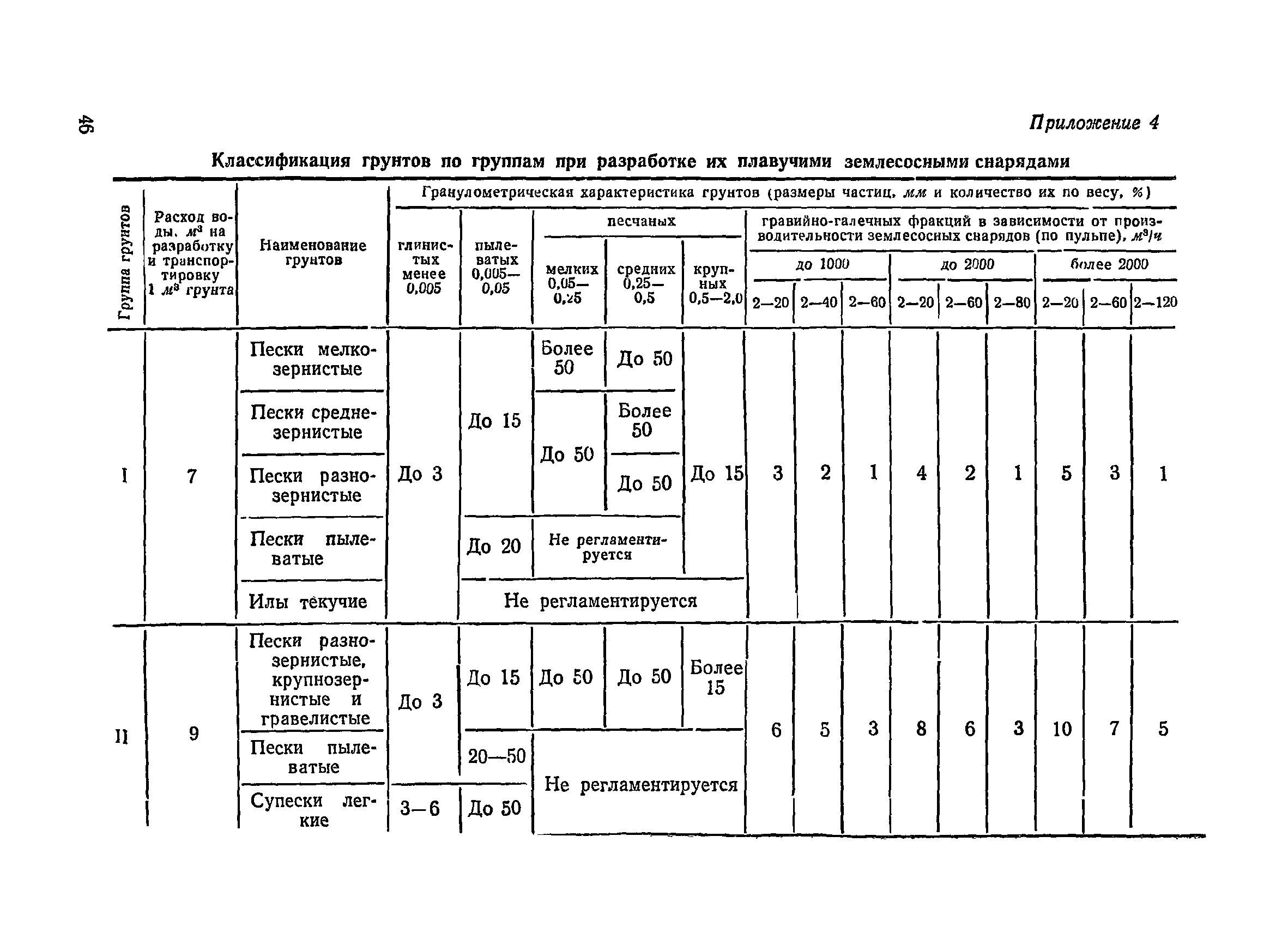 AtPIN2 определяет локус Arabidopsis для контроля корневого гравитропизма. EMBO J. 1998; 17: 6903–11.
AtPIN2 определяет локус Arabidopsis для контроля корневого гравитропизма. EMBO J. 1998; 17: 6903–11.
Артикул PubMed PubMed Central Google Scholar
Utsuno K, Shikanai T., Yamada Y, Hashimoto T. AGR, агравитропный локус Arabidopsis thaliana , кодирует новый член семейства мембранных белков.Physiol растительной клетки. 1998; 39: 1111–8.
Артикул CAS PubMed Google Scholar
Mravec J, Skůpa P, Bailly A, Hoyerová K, Bielach A, Petrášek J, et al. Субклеточный гомеостаз фитогормона ауксина опосредуется транспортером PIN5, локализованным в ER. Природа. 2009; 459: 1136–40.
Артикул CAS PubMed Google Scholar
Фримл Дж. Транспорт ауксина — формирование растения.Curr Opin Plant Biol. 2003; 6: 7–12.
Артикул CAS PubMed Google Scholar

Ding ZJ, Wang BJ, Moreno I., Dupláková N, Sibu S, Carraro N, et al. ER-локализованный транспортер ауксина PIN8 регулирует гомеостаз ауксина и развитие мужских гаметофитов у Arabidopsis. Nat Commun. 2012; 3: 941.
Артикул CAS PubMed Google Scholar
Rosquete MR, Waidmann S, Kleine-Vehn J.Носитель ауксина PIN7 играет преимущественную роль в прекращении радиального расширения корня у Arabidopsis thaliana . Int J Mol Sci. 2018; 19 (4): 1238.
Артикул CAS Google Scholar
Křeček P, Skůpa P, Libus J, Naramoto S, Tejos R, Friml J, Zažímalová E. Семейство белков-переносчиков ауксина, сформированное булавками (булавками). Genome Biol. 2009; 10: 249.
Артикул CAS PubMed PubMed Central Google Scholar
Рихтер С., Андерс Н., Вольтерс Х., Бекманн Х. , Томанн А., Генрих Р. и др. Роль гена GNOM в формировании апикально-базального паттерна Arabidopsis — от мутантного фенотипа до клеточного механизма действия белка. Eur J Cell Biol. 2010; 89: 138–44.
, Томанн А., Генрих Р. и др. Роль гена GNOM в формировании апикально-базального паттерна Arabidopsis — от мутантного фенотипа до клеточного механизма действия белка. Eur J Cell Biol. 2010; 89: 138–44.
Артикул CAS PubMed Google Scholar
Кохли П., Калия М., Гупта Р. Пектинметилестеразы: обзор. J Bioprocess Biotech. 2015; 5: 5.
Google Scholar
Ян XY, Zeng ZH, Yan JY, Fan W, Bian HW, Zhu MJ и др. Связь специфических пектинметилэстераз с ингибированием удлинения корней, индуцированным Al, у риса. Physiol Plant. 2013; 148: 502–11.
Артикул CAS PubMed Google Scholar
Daher BF, Chen Y, Bozorg B, Clough J, Jonsson H, Braybrook SA. Анизотропный рост достигается за счет аддитивного механического эффекта анизотропии материала и упругой асимметрии.Элиф. 2018; 7: e38161.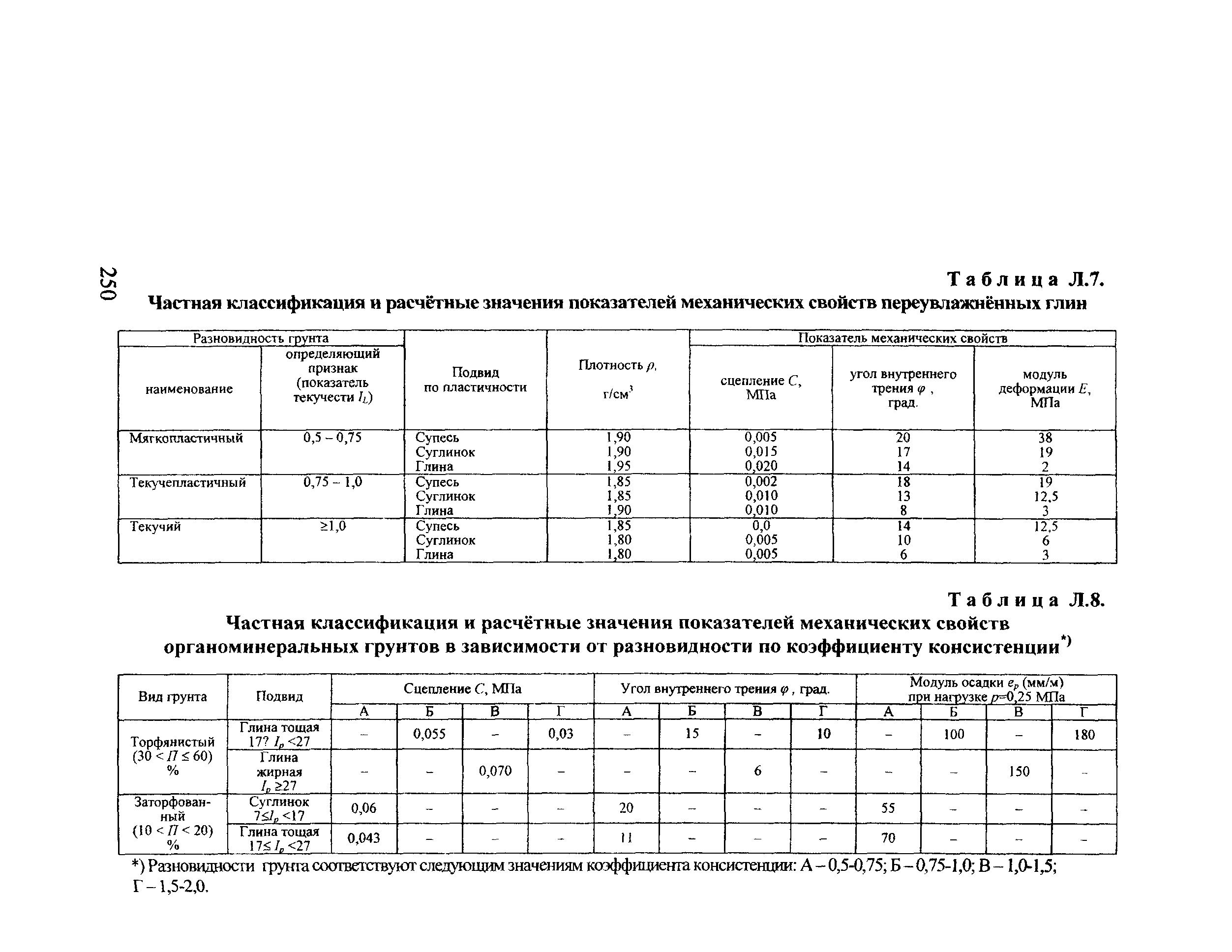
Артикул Google Scholar
Marchant A, Bhalerao R, Casimiro I, Eklöf J, Casero PJ, Bennett M, Sandberg G. AUX1 способствует формированию боковых корней, облегчая распределение индол-3-уксусной кислоты между раковиной и исходной тканью в проростке арабидопсиса . Растительная клетка. 2002. 14 (3): 589–97.
Артикул CAS PubMed PubMed Central Google Scholar
Chmielewska B, Janiak A, Karcz J, Guzy-Wrobelska J, Forster BP, Nawrot M, et al. Морфологические, генетические и молекулярные характеристики мутантов волосков корня ячменя. J Appl Genet. 2014; 55 (4): 433–47.
Артикул CAS PubMed PubMed Central Google Scholar
Инукай Ю., Сакамото Т., Уегучи-Танака М., Сибата Ю., Гоми К., Умемура И. и др. Crown rootless1, , который важен для образования корня кроны у риса, является мишенью AUXIN RESPONSE FACTOR в передаче сигналов ауксина. Растительная клетка. 2005; 17: 1387–96.
Растительная клетка. 2005; 17: 1387–96.
Артикул CAS PubMed PubMed Central Google Scholar
Liu H, Wang S, Yu X, Yu J, He X, Zhang S, Shou H, Wu P. ARL1, белок LOB-домена, необходимый для образования придаточных корней у риса. Плант Дж. 2005; 43 (1): 47–56.
Артикул CAS PubMed PubMed Central Google Scholar
Taramino G, Sauer M, Stauffer JL, Multani D, Niu X, Sakai H, Hochholdinger F.Ген RTCS кукурузы ( Zea mays L.) кодирует белок домена LOB, который является ключевым регулятором зародышевого семенного и постэмбрионального зародышевых корней. Плант Дж. 2007; 50: 649–59.
Артикул CAS PubMed PubMed Central Google Scholar
Роде В., Беккер Д., Саламини Ф. Структурный анализ восковидного локуса из Hordeum vulgare . Нуклеиновые кислоты рез. 1988; 16: 7185–718662. Тернер А., Билс Дж., Фор С., Данфорд Р.П., Д.а. Лори Д.А. Регулятор псевдоответа Ppd-h2 обеспечивает адаптацию к световому периоду ячменя. Наука. 2005. 310 (5750): 1031–4.
Артикул CAS Google Scholar
Douchkov D, Lück S, Johrde A, Nowara D, Himmelbach A, Rajaraman J, Stein N, Sharma R, Kilian B, Schweizer P. Открытие генов, влияющих на устойчивость ячменя к адаптированному и неадаптированному порошку грибки милдью. Genome Biol. 2014; 15 (12): 518.
Артикул CAS PubMed PubMed Central Google Scholar
Mikołajczak K, Ogrodowicz P, Gudyś K, Krystkowiak K, Sawikowska A, Frohmberg W, et al. Локусы количественных признаков для урожайности и связанных с урожайностью признаков в популяциях ярового ячменя, полученных от скрещивания европейских и сирийских сортов. PLoS One. 2016; 11 (5): e0155938.
Артикул CAS PubMed PubMed Central Google Scholar
Sharma R, Draicchio F, Bull H, Herzig P, Maurer A., Pillen K, Thomas W., Flavell AJ.Полногеномная ассоциация признаков урожайности в популяции ячменя, картирующей вложенные ассоциации, выявляет новое генетическое разнообразие для будущей селекции. Jounal Exp Bot. 2018; 69 (16): 3811–22.
Артикул CAS Google Scholar
Xu X, Sharma R, Tondelli A, Russell J, Comadran J, Schnaithmann F, et al. Полногеномный ассоциативный анализ признаков, связанных с урожайностью зерна, в панъевропейской коллекции сортов ячменя. Геном растений. 2018; 11 (1).https://doi.org/10.3835/plantgenome2017.08.0073.
Varshney RK, Gaur PM, ChamaRth SK, Krishnamurthy L, Tripathi S, Kashiwagi J, et al. Ускоренная интрогрессия «горячей точки QTL» по корневым признакам и другим признакам засухоустойчивости у JG 11, элитного и ведущего сорта нута. Геном растений. 2013; 6 (3): 1–9.
Google Scholar
Bull H, Casao MC, Zwirek M, Flavell AJ, Thomas WTB, Guo W. и др.Ячмень SIX-ROWED SPIKE3 кодирует предполагаемую деметилазу h4K9me2 / me3 Jumonji C-типа, которая подавляет фертильность латеральных колосков. Nat Commun. 2017; 8 (1): 1–9.
Артикул CAS Google Scholar
van Esse GW, Walla A, Finke A, Koornneef M, Pecinka A, von Korff M. Шестирядный колос 3 ( VRS3 ) представляет собой гистоновую деметилазу, которая контролирует развитие латеральных колосков ячменя. Plant Physiol. 2017; 174: 2397–408.
Артикул CAS PubMed PubMed Central Google Scholar
Liller CB, Neuhaus R, Von Korff M, Koornneef M, Van Esse W. Мутации в генах ячменного ряда оказывают плейотропное воздействие на ветвление побегов. PLoS One. 2015; 10 (10): 1–20.
Артикул CAS Google Scholar
Werner T, Motyka V, Laucou V, Smets R, Van Onckelen H, Schmülling T. Трансгенные растения Arabidopsis с дефицитом цитокинина демонстрируют множественные изменения в развитии, указывающие на противоположные функции цитокининов в регуляции активности меристем побегов и корней.Растительная клетка. 2003. 15 (11): 2532–50.
Артикул CAS PubMed PubMed Central Google Scholar
Osugi A, Sakakibara HQ. A: как растения реагируют на цитокинины и в чем их важность? BMC Biol. 2015; 13: 102.
Артикул CAS PubMed PubMed Central Google Scholar
Alqudah AM, Koppolu R, Wolde GM, Graner A, Schnurbusch T.Генетическая архитектура роста растений ячменя. Фронт Жене. 2016; 7: 117.
Артикул CAS PubMed PubMed Central Google Scholar
Маурер А., Драба В., Цзян Ю., Шнайтманн Ф., Шарма Р., Шуман Э. и др. Моделирование генетической архитектуры контроля времени цветения ячменя посредством картирования вложенных ассоциаций. BMC Genomics. 2015; 16: 290.
Артикул CAS PubMed PubMed Central Google Scholar
Маурер А., Драба В., Пиллен К. Геномное расчленение развития растений и его влияние на массу тысячи зерен ячменя посредством картирования вложенных ассоциаций. J Exp Bot. 2016; 67 (8): 2507–18.
Артикул CAS PubMed PubMed Central Google Scholar
Ingvordsen CH, Backes G, Lyngkjr MF, Peltonen-Sainio P, Jahoor A, Mikkelsen TN, Jørgensen RB. Полногеномное ассоциативное исследование продуктивности и характеристик устойчивости ячменя, культивируемого при будущих климатических сценариях.Мол Порода. 2015; 35: 84.
Артикул CAS Google Scholar
Саннеманн В., Хуанг Б.Э., Мэтью Б., Леон Дж. Скрещивание многоплодных передовых поколений ячменя: количественное картирование локусов признаков с высоким разрешением для времени цветения в качестве доказательства концепции. Мол Порода. 2015; 35: 86.
Артикул CAS Google Scholar
Alqudah AM, Youssef HM, Graner A, Schnurbusch T.Естественная изменчивость и генетический состав площади листовой пластинки ярового ячменя. Theor Appl Genet. 2018; 131: 873–86.
Артикул CAS PubMed PubMed Central Google Scholar
Тернер А., Билс Дж., Фор С., Данфорд Р.П., Лори Д.А. DA. Регулятор псевдоответа Ppd-h2 обеспечивает адаптацию к световому периоду ячменя. Наука. 2005. 310 (5750): 1031–4.
Артикул CAS PubMed Google Scholar
Бейкер К., Байер М., Кук Н., Дрессиг С., Диллон Т., Рассел Дж. И др. Перицентромерная область ячменя с низким уровнем рекомбинации ограничивает разнообразие и эволюцию генов, но не экспрессию генов. Плант Дж. 2014; 79: 981–92.
Артикул CAS PubMed PubMed Central Google Scholar
Helariutta Y, Fukaki H, Wysocka-Diller J, Nakajima K, Jung J, Sena G, Hauser MT, Benfey PN. Ген SHORT-ROOT контролирует формирование радиального паттерна корня Arabidopsis посредством радиальной передачи сигналов.Клетка. 2000. 101 (5): 555–67.
Артикул CAS PubMed PubMed Central Google Scholar
Benitez-Alfonso Y, Jackson D. Редокс-гомеостаз регулирует плазмодесмальные коммуникации в меристемах Arabidopsis. Сигнальное поведение растений. 2009. 4 (7): 655–9.
Артикул CAS PubMed PubMed Central Google Scholar
Веласкес С.М., Рикарди М.М., Поулсен С.П., Оикава А., Дилокпимол А., Халим А. и др.Комплексная регуляция пролил-4-гидроксилазы влияет на рост корневых волос. Завод Мол. 2015; 8 (5): 734–46.
Артикул CAS PubMed PubMed Central Google Scholar
Сакума С., Пуркхейрандиш М., Хенсель Г., Кумлен Дж., Стейн Н., Тагири А. и др. Дивергенция паттерна экспрессии способствовала неофункционализации дублированного транскрипционного фактора HD-zip I в ячмене. Новый Фитол. 2013; 197 (3): 939–48.
Артикул CAS PubMed PubMed Central Google Scholar
de la Rosa Santamaria R, Shao MR, Wang G, Nino-Liu DO, Kundariya H, et al. Негенетическая изменчивость, индуцированная MSh2, обеспечивает источник фенотипического разнообразия у Sorghum bicolor. PLoS One. 2014; 9 (10): e108407.
Артикул CAS PubMed PubMed Central Google Scholar
Вирди К.С., Лори Дж. Д., Сюй И-Зи, Ю Дж., Шао М.-Р., Санчес Р. и др. Мутация MSh2 Arabidopsis изменяет эпигеном, вызывая наследственные изменения в росте растений.Nat Commun. 2015; 6: 6386.
Артикул CAS PubMed PubMed Central Google Scholar
Вирди К.С., Вамболдт Й., Кундария Х., Лори Д.Д., Керен И., Кумар К.С., Блок А и др. MSh2 — это связывающий органеллярную ДНК растений и тилакоидный белок, находящийся под точной пространственной регуляцией для изменения развития. Завод Мол. 2016; 9: 245–60.
Артикул CAS PubMed PubMed Central Google Scholar
Наир С.К., Ван Н., Туруспеков Ю., Пуркхейрандиш М., Синсувонгват С., Чен Г. и др. Клейстогамное цветение ячменя возникает в результате подавления регулируемого микроРНК расщепления мРНК HvAP2. Proc Natl Acad Sci. 2010. 107 (1): 490–5.
Артикул PubMed PubMed Central Google Scholar
Хьюстон К., МакКим С.М., Комадран Дж., Бонар Н., Друка И., Узрек Н. и др. Вариация взаимодействия аллелей HvAPETALA2 и микроРНК172 определяет плотность зерен на соцветиях ячменя.Proc Natl Acad Sci. 2013. 110 (41): 16675–80.
Артикул PubMed PubMed Central Google Scholar
Licausi F, Ohme-Takagi M, Perata P. Факторы транскрипции APETALA2 / этилен-чувствительный фактор (AP2 / ERF): медиаторы стрессовых реакций и программ развития. Новый Фитол. 2013; 199 (3): 639–49.
Артикул CAS PubMed PubMed Central Google Scholar
Рихтер Дж., Плодерер М., Монгелард Дж., Гутьеррес Л., Хаузер М.-Т. Роль CrRLK1L датчиков клеточной стенки HERCULES1 и 2, THESEUS1 и FERONIA в адаптации роста, вызванной тяжелыми металлами и микроэлементами. Фронтальный завод им. 2017; 8: 1554.
Артикул PubMed PubMed Central Google Scholar
Schwartz SH, Qin X, Zeevaart JAD. Выяснение непрямого пути биосинтеза абсцизовой кислоты мутантами. Гены и ферменты Plant Physiol.2003. 131 (4): 1591–601.
Артикул CAS PubMed PubMed Central Google Scholar
Harris JM. Абсцизовая кислота: скрытый архитектор структуры корневой системы. Растения (Базель). 2015; 4 (3): 548–72.
Артикул CAS Google Scholar
Sharp RE, LeNoble ME. АБК, этилен и контроль роста побегов и корней в условиях водного стресса. J Exp Bot.2002. 53 (366): 33–7.
Артикул CAS PubMed PubMed Central Google Scholar
Rollins JA, Drosse B, Mulki MA, Grando S, Baum M, Singh M, S. Ceccarelli A, von Korff M. Вариация генов яровизации Vrn-h2 и Vrn-h3 определяет стабильность роста и урожайности ячменя ( Hordeum vulgare ), выращенного в условиях засушливых земель в Сирии. Теор Аппл Генет, 2013; 126 (11): 2803–2824.
Griffiths S, Dunford RP, Coupland G, Laurie DA. Эволюция семейств генов, подобных CONSTANS , у ячменя, риса и Arabidopsis. Plant Physiol. 2003. 131 (4): 1855–67.
Артикул CAS PubMed PubMed Central Google Scholar
Кокрам Дж., Тиль Т., Штойернагель Б., Стейн Н., Таудьен С., Бейли П.С., О’Салливан Д.М. Динамика генома объясняет эволюцию семейств генов домена CCT во время цветения у Poaceae.PLoS One. 2012; 7 (9): e45307.
Артикул CAS PubMed PubMed Central Google Scholar
Campoli C, Drosse B, Searle I, Coupland G, Von Korff M. Функциональная характеристика HvCO1, ячменя ( Hordeum vulgare ), время цветения, ортолога CONSTANS. Плант Дж. 2012; 69 (5): 868–80.
Артикул CAS PubMed PubMed Central Google Scholar
Calixto CPG, Simpson CG, Waugh R, Brown JWS. Альтернативный сплайсинг часовых генов ячменя в ответ на низкую температуру. PLoS One. 2016; 11 (12): e0168028.
Артикул CAS PubMed PubMed Central Google Scholar
Li XY, Qian Q, Fu ZM, Wang YH, Xiong GS, et al. Контроль кущения риса. Природа. 2003; 422: 618–21.
Артикул CAS PubMed PubMed Central Google Scholar
Zhang XH, Rao XL, Shi HT, Li RJ, Lu YT. Сверхэкспрессия гена цитозольной глицеральдегид-3-фосфатдегидрогеназы OsGAPC3 придает рису солеустойчивость. Культ растительных клеток, тканей и органов. 2011; 107: 1–11.
Артикул CAS Google Scholar
Ноч Э, Дилокпимол А, Трифона Т., Поульсен С.П., Сюн Дж., Хархольт Дж. И др. Бета-глюкуронозилтрансфераза из Arabidopsis thaliana , участвующая в биосинтезе арабиногалактана типа II, играет роль в удлинении клеток во время роста проростков.Плант Дж. 2013; 76: 1016–29.
Артикул CAS PubMed PubMed Central Google Scholar
Го М., Лю Дж.Х., Ма Х, Ло Д-Х, Гун З. Х., Лу М-Х. Факторы транскрипции теплового стресса (HSF) растений: структура, регуляция и функция в ответ на абиотические стрессы. Фронтальный завод им. 2016; 7: 114.
PubMed PubMed Central Google Scholar
Осакабе И, Аринага Н., Умедзава Т., Кацура С., Нагамати К., Танака Х., Охираки Х., Ямада К., Сео С.-У, Або М. и др. Осмотические реакции на стресс и рост растений, контролируемые переносчиками калия в Arabidopsis . Растительная клетка. 2013; 25: 609–24.
Артикул CAS PubMed PubMed Central Google Scholar
Грейтер С.П., Боддингтон К.Ф. Нарушение и функция: обзор семейства белков дегидрина. Фронтальный завод им.2014; 5: 576.
Артикул PubMed PubMed Central Google Scholar
Trevaskis B, Tadege M, Hemming MN, Peacock WJ, Dennis ES, Sheldon C. Гены MADS-бокса, подобные короткой вегетативной фазе, ингибируют идентичность цветочной меристемы ячменя. Plant Physiol. 2007. 143: 225–35.
Артикул CAS PubMed PubMed Central Google Scholar
Ким С.Ю.Роль факторов транскрипции класса bZIP семейства ABF в стрессовой реакции. Physiol Plant. 2006; 126: 519–27.
CAS Google Scholar
Uno Y, Furihata T., Abe H, Yoshida R, Shinozaki K, Yamaguchi-Shinozaki K. Основные факторы транскрипции лейциновой молнии Arabidopsis, участвующие в пути передачи сигнала, зависимого от абсцизовой кислоты, в условиях засухи и высокой солености. Proc Natl Acad Sci U S. A. 2000; 97: 11632–7.
Артикул CAS PubMed PubMed Central Google Scholar
Ким С., Кан Дж.Й., Чо Ди, Пак Дж. Х., Ким С. ABF2, ABRE-связывающий фактор bZIP, является важным компонентом передачи сигналов глюкозы, и его сверхэкспрессия влияет на устойчивость к множественному стрессу. Плант Дж. 2004; 40 (1): 75–87.
Артикул CAS PubMed PubMed Central Google Scholar
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Проектирование земляных работ | FHWA
Следующая информация дополняет раздел 9 PDDM.5.1.
В то время как выемка проезжей части не классифицируется для целей измерения или оплаты, ее можно разделить на следующие категории для расчета проектных и массовых расчетов земляных работ:
- Обычный материал . Обычный материал — это в основном земля или земля с отдельными валунами менее 0,5 куб.
- Рыхлая порода . Рыхлая порода — это материал, готовый к выемке после разрыхления рыхлителем.
- Солид Рок .Твердая порода включает твердые породы на месте, уступы и валуны, для удаления которых требуется буровзрывное оборудование. Любые взрывные работы будут проводиться в соответствии с техническими условиями взрывного участка.
Определение объемов земляных работ и насыпи
Используйте метод средней конечной площади для определения объемов. Общий объем земляных работ складывается из объемов призмоидов, образованных смежными поперечными сечениями.
При использовании метода средней концевой площади призмоид рассматривается как призма, поперечное сечение которой является средним из двух концевых площадей призмоида.Уравнение 9.5.1A (1) представляет собой формулу для использования в методе средней конечной площади.
Уравнение 9.5.1A (1)
Где:
V = Объем, кубический ярд [м 3 ]
A 1 и A 2 = Концевые площади поперечного сечения, квадратных футов [м 2 ]
L = Расстояние между поперечными сечениями, фут [м]
Эта формула приблизительно верна. Благодаря своей простоте и значительной точности в большинстве случаев она стала широко используемой формулой.Это дает результаты, в целом, превышающие истинный объем.
Когда центр масс земляных работ (центроид области выемки или насыпи) не отцентрован относительно проезжей части, а трасса находится в кривизне, фактический вычисленный объем неверен, поскольку истинное расстояние между центроидами конечной области будет отличаться с расстояния по средней линии. В этом случае может потребоваться отрегулировать объемы выемки грунта с учетом кривизны, чтобы правильно учесть земляные работы. Во многих случаях в этом нет необходимости, поскольку эксцентриситет относительно осевой линии массы земляных работ имеет тенденцию выравниваться по маршруту.
Коэффициенты усадки и набухания
Используя данные, предоставленные инженерно-геологическим отделом, проектировщик должен проверить характеристики материала, который будет выкапывать или укладывать в насыпи. Выемка, используемая для строительства насыпей, будет варьироваться от скальных пород до земли и будет иметь коэффициенты усадки / набухания, назначенные для целей проектирования.
Значения, показанные в Приложении 5.1A, могут использоваться для целей оценки до получения информации о конкретном проекте от Геотехнического подразделения.
| Материал | Измерено | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| In-situ Массовая плотность 1 | Насыпь | |||||||||||||||||||||||||
| Массовая плотность 2 | % Swell 3 | |||||||||||||||||||||||||
| фунт / фут 3 | кг / м 3 | фунт / фут 3 | 922 м 3фунт / фут 3 | кг / м 3 | ||||||||||||||||||||||
| Андезит | 4950 | 2930 | 2970 | 1760 | 67 | 1760 | 67 | 4 344 9234 9234|||||||||||||||||||
| Базальт | 4950 | 2935 | 3020 | 1790 | 64 | 3640 | 2160 | 36 | 9 2284||||||||||||||||||
| Бентонит | 2700 | 1600 | 2000 | 1185 | 35 | — | — | — | ||||||||||||||||||
| Брекчия | 4050343 | |||||||||||||||||||||||||
| 9234 9234 9234 9234 9234 9234 | 3190 | 1890 | 27 | |||||||||||||||||||||||
| Кальцит-кальций | 4500 | 2670 | 2700 | 1600 | 67 | — | — | — | 93432100 | 1245 | 16 | 3200 | 1900 | -25 | ||||||||||||
| Мел | 4060 | 2410 | 2170 | 12223 | 2170 | 9344 | Древесный уголь | — | — | 1030 | 610 | — | — | — | — | |||||||||||
| Пепели | 1280 | 760 | 960 | 570 | 33 | 1420 | 840 | -10 | 840 | -10 | 840 | -10 | 4 8 92348 9220 92348 1910 1985 | 2150 2010 | 1275 1180 | 50 67 | 35702248 3720 | 022034 0248 220349 0248 220348 Конгломерат | 3720 | 2205 | 2800 | 1660 | 33 | — | — | — |
| Разложившаяся порода — 75% R.25% E. — 50% Р. 50% E. — 25% Р. 75% E. | 4120 3750 3380 | 2445 2225 2005 | 3140 2710 2370 | 1865 1610 1405 | 318248 43248 2185 2375 2205 | 12 -6 -9 | ||||||||||||||||||||
| Диорит | 5220 | 3095 | 3130 | 1855 | 67 | 9343 1855 | 67 | 9343 | земля | 1470 | 870 | 910 | 540 | 62 | — | — | — | |||||||||
| Доломит | 4870 | 28903 17343 | 9234 9234 92342015 | 43 | ||||||||||||||||||||||
| Земля, суглинок — Сухая — Влажная — Влажная, грязь | 3030 3370 2940 | 1795 2000 1745 | 2070 2360 2940 | 1230 1400 1745 | 50 43 0 | -12 -4 -20 | ||||||||||||||||||||
| Полевой шпат | 4410 | 2615 | 2640 | 1565 | 67 | 3080 9225 434344 9343 | 9225 9344 9343 | 9344 | | 3095 | 3130 | 1855 | 67 | 3650 | 2165 | 43 | ||||||||||
| Гнейс | 4550 | 2700 | 9234 9234 9234 9234 9234 9234 9234 9234 9234 9234 9234 9234 9234 | 92349234 9234 9234 9234 9234 9234 9234 9234 9234 9234 | 9234 9234 9234 9234 9234 9234 9234 9234 9234 9234 9234 9234||||||||||||||||||||||
| Гравий (сухой) — Однородный — Средн.Градация — Хорошая градация | 2980 3280 3680 | 1770 1945 2180 | 2700 2730 2770 | 1600 1620 | 1870 2120 2450 | -5 -8 -11 | ||||||||||||||||||||
| Гравий (мокрый) — равномерно гранулированный — средн. Градация — Ну Graded | 3310 3640 4090 | 1965 2160 2425 | 3150 3290 3520 | 1870 1950 2090 | 5 10 16 | 3150 3570 4130 | 1870 2120 2450 | -5 -2 -1 | ||||||||||||||||||
| Гранит | 4540 | 2695 | 2640 | 153 | 9234 9234 9234 9234 9234 43||||||||||||||||||||||
| Гамбо — Сухой — Мокрый | 3230 3350 | 1915 1985 | 2150 2020 | 1275 1200 | 1275 1200 | 1275 1200 | -10 -10 | |||||||||||||||||||
| Гипс | 4080 | 2420 | 2380 92 344 | 1410 | 72 | — | — | — | ||||||||||||||||||
| Магматические породы | 4710 | 2795 | 2820 | 1675 | 67 | 9344 9344 9234 9234 9234 9234 9234 9234 9234 9234 9234 9234 9234 9234 9234 9344 — Сухой — Мокрый | 3230 3350 | 1915 1985 | 2150 2010 | 1275 1190 | 50 67 | | ||||||||||||||
| Известняк | 4380 | 2600 | 2690 | 1595 | 63 | 3220 | 1910 | 36 | 1910 | 36 | ||||||||||||||||
| 1910 1985 | 2150 2010 | 1275 1190 | 50 67 9 2344 | 3570 3720 | 2120 2205 | -10 -10 | ||||||||||||||||||||
| Мрамор | 4520 | 2680 | 2700 | 1600 | 9234 9234 9234 9234 9234 9234||||||||||||||||||||||
| Марл | 3740 | 2220 | 2240 | 1330 | 67 | 2620 | 1555 | 43 | ||||||||||||||||||
| Кладка, щебень 44 | ||||||||||||||||||||||||||
| 9234 9234 9234 9234 9234 9234 9234 9234 | 2750 | 1630 | 43 | |||||||||||||||||||||||
| Слюда | 4860 | 2885 | 2910 | 1725 | 67 | — | — 92avement4 | 92828 — Бетон 3240 4050 3960 2840 | 1920 2400 9224 8 2350 1685 | 1940 2430 2370 1700 | 1150 1440 1405 1010 | 50 67 67 67 | 27348 27 | 9240 2734 2834 2734 2834 1645 1685 | 0 43 43 0 | |||||||||||
| Торф | 1180 | 700 | 890 | 530 | 33 | — 44 4 | 4 | 4 | 4 | 92 1080640 | 650 | 385 | 67 | — | — | — | ||||||||||
| Кварц | 4360 | 2585 | 4360 | 2585 | 2610 | 9234 9234 9234 9234 | 2610 | 158592 43 | ||||||||||||||||||
| Кварцит | 4520 | 2680 | 2710 | 1610 | 67 | 3160 | 92 343 187543 | |||||||||||||||||||
| Риолит | 4050 | 2400 | 2420 | 1435 | 67 | 2870 | 1700 | 4210 | 43 | 99 | 9999 | 4 | 1550 | 72 | 3150 | 1870 | 43 | |||||||||
| Песок — Сухой — Мокрый | 2880 3090 | 1710 1915 34 | 1710 1915 34 34 | 11 5 | 3240 3460 | 1920 2050 | -11 -11 | |||||||||||||||||||
| Песчаник | 4070 | 2415 | 4070 | 2415 | 25203 1795 | 34 | ||||||||||||||||||||
| Сланец | 4530 | 2685 | 2710 | 161 0 | 67 | 3170 | 1880 | 43 | ||||||||||||||||||
| Сланец | 4450 | 2640 | 2480 | 1470 | 79 | 2990 1770 | 2990 97534 9344 9234 9234 9234 9234 99234 9234 9234 99234 | 1920 | 2380 | 1410 | 36 | 3890 | 2310 | -17 | ||||||||||||
| Алевролит | 4070 | 2415 | 1495344 | 2415 | 25203 1495344 | 2415 | 25203 1495344 | 2415 | 25203 1495344 | 2415 | 25203 1495344 | 2415 | 25203 1495344 -11 | |||||||||||||
| Сланец | 4500 | 2670 | 2600 | 1540 | 77 | 3150 | 1870 | 43 | ||||||||||||||||||
| 43 | 3250 | 1930 | 43 | |||||||||||||||||||||||
| Верхний слой почвы | 2430 | 9 2343 14401620 | 960 | 56 | 3280 | 1945 | -26 | |||||||||||||||||||
| Туф | 4050 | 1820 | 4050 | 2400 | 2700 | 1600 | 1600 | 1600 | 1600 | |||||||||||||||||
Сноски:
- Возможны отклонения в среднем ± 5%.
- Массовая плотность может быть скорректирована в соответствии с модифицированными коэффициентами набухания и усадки.
- На основе средней плотности на месте. Отрицательное число означает усадку. Факторы, допускающие отклонение ± 33%.
выемка проезжей части обычно измеряется в исходном, ненарушенном положении. В спецификациях должны четко указываться место и метод измерения, потому что почти все материалы изменяют объем при движении от пропила к насыпи.
Извлеченный обычный материал будет расширяться за пределы своего первоначального объема в транспортном средстве, но, как правило, сжимается ниже вынутого объема при уплотнении в насыпь.Для иллюстрации, 1 куб. Д. [1 м3] земли в разрезе может занимать 1,25 куб. Д. [1,25 м3] пространства в транспортном средстве и, наконец, занимать только 0,65–0,85 куб. Это, конечно, зависит от его исходной плотности и приложенного усилия уплотнения. Эта разница между исходным объемом в разрезе и окончательным объемом в заливке и есть усадка.
Выемка твердой породы, помещенная в насыпь, обычно занимает больший объем. Это изменение объема и есть зыбь. Когда пустоты в каменной насыпи заполняются землей или другим мелким материалом, объем засыпки будет примерно равен объединенным объемам в двух местах расположения источников.
При выемке легких грунтов и насыпях, сооружаемых на болотах, подверженных осадкам, усадка может составлять от 20 до 40 процентов или даже больше. При умеренной выемке грунта усадка составляет от десяти до 25 процентов. При выемке тяжелых грунтов с глубокими выемками и насыпями следует ожидать усадки от примерно 15 процентов до набухания в пять процентов. Усадка обычно включает незначительные потери при транспортировке материала от пропила к насыпи и потерю материала, выходящего за пределы склонов.Набережные, которые слегка застроены, также способствуют явной усадке.
Нижележащий грунт под насыпью также может оседать и уплотняться или смещаться из-за размещения и уплотнения насыпи, что приводит к явной усадке. Оседание приводит к очевидной усадке, но одно не прямо пропорционально другому. Не путайте усадку с проседанием. Просадка — это оседание всей насыпи из-за слабых условий фундамента (например, укладка насыпи на заболоченный грунт).
При выемке породы часто ожидается набухание от 5 до 25 процентов в зависимости от доли твердой породы и размера породы, помещенной в насыпь. Обработка горных пород, особенно взрывных работ с подушками, является неточной и может привести к небольшому излишку выемки на откосах, что приведет к явному вздутию.
Если возможно, при проектировании следует учитывать фактические факторы усадки и набухания в полевых условиях для аналогичного материала, используемого в смежных проектах.
Балансировка земляных работ
Часто бывает, что материала из соседних пропилов недостаточно для заполнения промежуточной насыпи.В этом случае материал заимствуется из-за пределов строительства.
При избытке выкопанного материала может потребоваться его утилизация. Вместо длительных перевозок может быть более экономичным утилизировать материал, расширив обочины или разместив материал на площадках для захоронения, чем оплачивать затраты на транспортировку.
Если земляные работы не сбалансированы, проектировщик должен попытаться отрегулировать линию уклона или осевую линию так, чтобы они были сбалансированы. Когда сбалансированный проект нецелесообразен или нежелателен, дизайнер либо избавляется от лишнего материала, либо заимствует материал для достижения баланса.Для выделенных участков захоронения или займов требуется разрешение на надлежащее владение, права пользования, экологические соображения и соответствующие разрешения.
Отходы для размещения излишков материала и / или участки карьера должны быть указаны на планах.
Транспортировка
Транспортировка состоит из транспортировки материала от его исходного положения к его конечному местоположению. Стоимость перевозки материала необходима для оценки удельной стоимости различных работ.