Типовые узлы — Изобент
Применение бентонитовой гидроизоляции
Схема укладки бентонитовых матов под фундаментной плитой
Схема перехода горизонтальной гидроизоляции на вертикальную поверхность
Технологический промежуток бетонирования фундаментной плиты
Переход горизонтальной изоляции на плиту фундамента в случае плиты с выступом
Узел гидроизоляции в случае, когда плита пола выше толщи фундамента
Переход горизонтальной гидроизоляции на вертикальную поверхность с использованием бентонитовой шпатели
Установка бентонитовых матов под температурный швом в фундаментальной плите (с иполльзованием внешней гидрошпонки)
Установка бентонитовых матов под температурным швом в фундаментной плите (с использованием внутренней гидрошпонки)
Гидроизоляция балки усиления фундамента
Изоляция балки усиления фундамента
Устройство гидроизоляции перехода инженерной коммуникации
Устройство гидроизоляции группы переходов инженерной коммуникации
Схема уплотнения отверстия после снятия опалубки
Крепление мата ИЗОБЕНТ по вертикали
Узлы крепления матов к стене фундамента ниже уровня грунта
Изоляция нижней части приямка шахт лифтов и других заглубленных конструкций
Изоляция нижней части приямка шахт лифтов и других заглубленных конструкций
Деталь гидроизоляции в зоне шахты лифта
Гидроизоляция фундамента
Схема гидроизоляции свайного фундамента
Изоляция нижней части приямка шахт лифтов и других заглубленных конструкций
Система слоев изоляции нижней части полигона
Важные моменты и узлы при устройстве столбчатого фундамента.

Как избежать распространённых ошибок при закладке столбчатого фундамента при соблюдении основных принципов технологического процесса.
● Возведённые на пучинистых грунтах фундаменты и оставленные на зиму без нагрузки — то есть без стен, перекрытий и крыш, могут подвергнуться деформации. Поэтому строительство дома на таких типах грунтов необходимо производить в один строительный сезон. Для столбчатых фундаментов этот момент особенно актуален ввиду того, что по причине отсутствия единого жёсткого остова каждый столб по своей сути сам является отдельным фундаментом. При этом каждый столб даёт свою собственную осадку и в дальнейшем пр устройстве ростверка и коробки строения могут возникнуть серьёзные сложности.
● Для начала фундаментных работ наиболее благоприятным
периодом времени считается время, когда почва отошла от зимней стужи и
грунтовые воды ушли на нижние пласты. В средней полосе России к этому
времени относятся летний период и начало осени. Если уже возведённое в
зимний период строение не эксплуатируется и не отапливается, а глубина
заложения фундамента была рассчитана на отапливаемый режим эксплуатации,
также могут произойти «нежданные» деформации.
В средней полосе России к этому
времени относятся летний период и начало осени. Если уже возведённое в
зимний период строение не эксплуатируется и не отапливается, а глубина
заложения фундамента была рассчитана на отапливаемый режим эксплуатации,
также могут произойти «нежданные» деформации.
● Если опоры столбчатого фундамента выполнены из монолитного бетона, то не стоит забывать тот факт, что потребуется 28 дней для достижения бетоном своих рабочих характеристик. За этот срок нельзя подвергать бетон каким-либо нагрузкам и уберечь бетонные столбы от негативных внешних погодных факторов — укрыть рубероидом или плёнкой и в зависимости от погодных условий несколько раз в неделю смачивать водой для равномерного схватывания.
● Приготовлением бетона для столбчатого фундамента лучше
заниматься с применением
цемента М400. В качестве наполнителя подходит песок с крупными зёрнами и
мелкий гравий.
● При самостоятельных работах во время приготовления бетонной смеси необходимо соблюдать все требования пропорциональности компонентов. Слишком густой или излишне жидкий бетон могут привести к уменьшению прочности бетонной конструкции до 25%
| Основные ошибки при заложении фундамента |
● К основным дефектам столбчатого фундамента относится неравномерность его проседания. Этот негативный результат появляется по следующим причинам:
• Фундаментные опоры имеют разную величину заглубления, расчёт глубины
закладки был выполнен с ошибками, неравномерная нагрузка на фундаментные
опоры. Для правильного расчёта нагрузки необходимо выполнить достаточно
точный расчёт планомерного распределения нагрузки с учётом всевозможных
надстроек.
Для правильного расчёта нагрузки необходимо выполнить достаточно
точный расчёт планомерного распределения нагрузки с учётом всевозможных
надстроек.
• Оценка несущих свойств грунта была произведена неправильно.
• Использование материала не надлежащего качества. К примеру, после хранения цемента полгода его марка снижается на 25%, а хранение около года и более — на 35-50%. Применение содержащего глину песка во время приготовления бетонной смеси также снижает характеристики бетона.
| Узлы столбчатого фундамента |
Расчет состава и оценка трещинообразования бетона в раннем возрасте
Аннотация:
В разных городских районах Вьетнама построено много крупномасштабных сооружений из монолитных бетонных конструкций.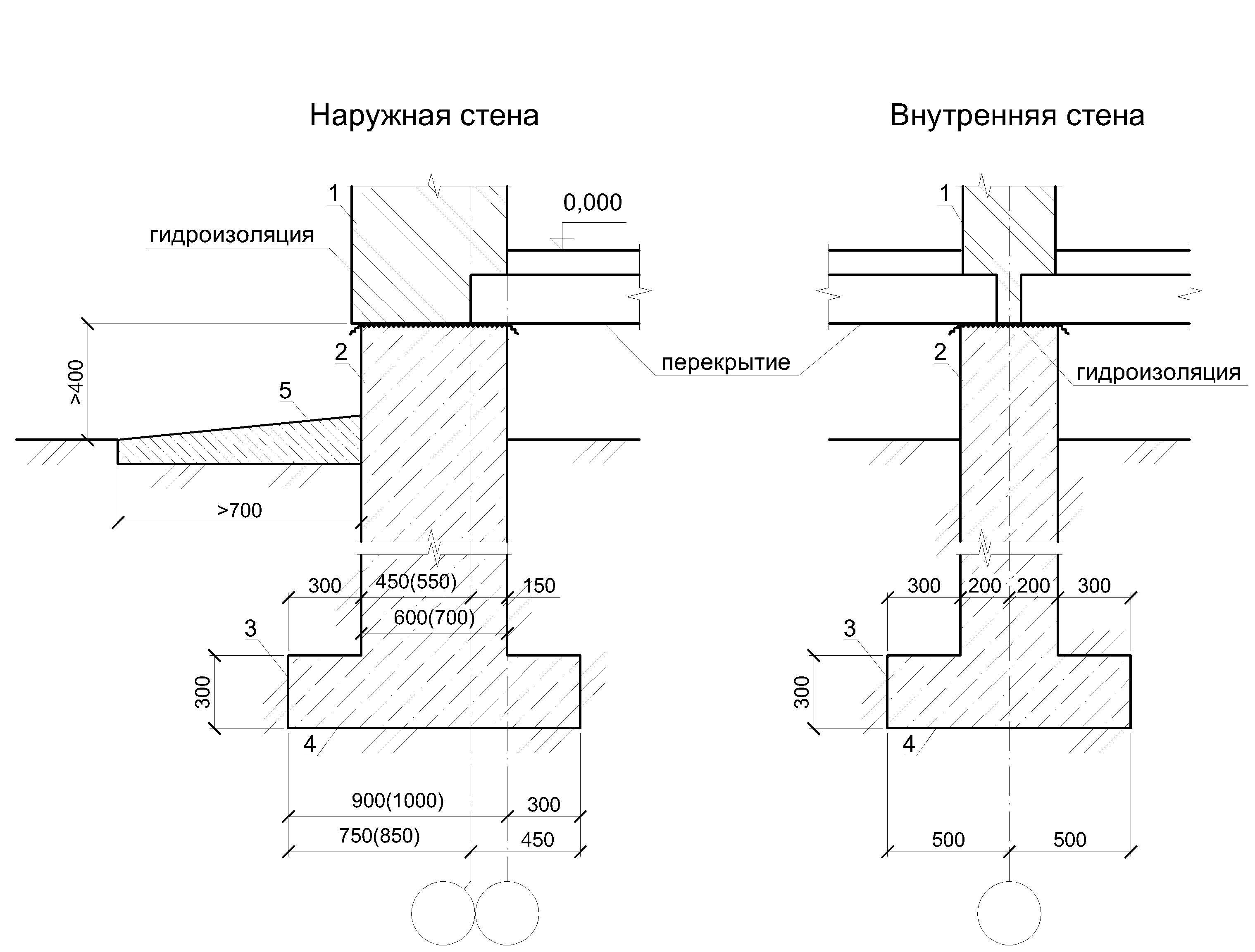 Однако, повреждение и растрескивание конструкций, вызванное температурными напряжениями, становятся все более распространенными и сильно сказываются на их надежности и долговечности эксплуатации. В работе для определения состава тяжёлого бетона для строительства фундамента моста был использован американский стандарт ACI 211.1-02. Оценка возможности трещинообразования в бетонном фундаменте в раннем возрасте была выполнена путём анализа температурного режима и возникающего в нём термонапряжения. В результате проведённых исследований была доказана возможность получения тяжёлого бетона из местных сырьевых материалов Вьетнама, с удобоукладываемостью бетонной смеси по осадке стандартного конуса 95 мм, обладающего прочностью на сжатие 36,3 МПа в возрасте 28 суток нормального твердения и средней водонепроницаемостью серии образцов 0,32 МПа для строительства моста фундамента размером 8 х 6 х 2,5 м. С помощью компьютерной программы MIDAS CIVIL была определена максимальная температура в центре бетонного фундамента по истечении 72 часов с момента затворения водой, равная Тmax = 73,04 °С.
Однако, повреждение и растрескивание конструкций, вызванное температурными напряжениями, становятся все более распространенными и сильно сказываются на их надежности и долговечности эксплуатации. В работе для определения состава тяжёлого бетона для строительства фундамента моста был использован американский стандарт ACI 211.1-02. Оценка возможности трещинообразования в бетонном фундаменте в раннем возрасте была выполнена путём анализа температурного режима и возникающего в нём термонапряжения. В результате проведённых исследований была доказана возможность получения тяжёлого бетона из местных сырьевых материалов Вьетнама, с удобоукладываемостью бетонной смеси по осадке стандартного конуса 95 мм, обладающего прочностью на сжатие 36,3 МПа в возрасте 28 суток нормального твердения и средней водонепроницаемостью серии образцов 0,32 МПа для строительства моста фундамента размером 8 х 6 х 2,5 м. С помощью компьютерной программы MIDAS CIVIL была определена максимальная температура в центре бетонного фундамента по истечении 72 часов с момента затворения водой, равная Тmax = 73,04 °С.
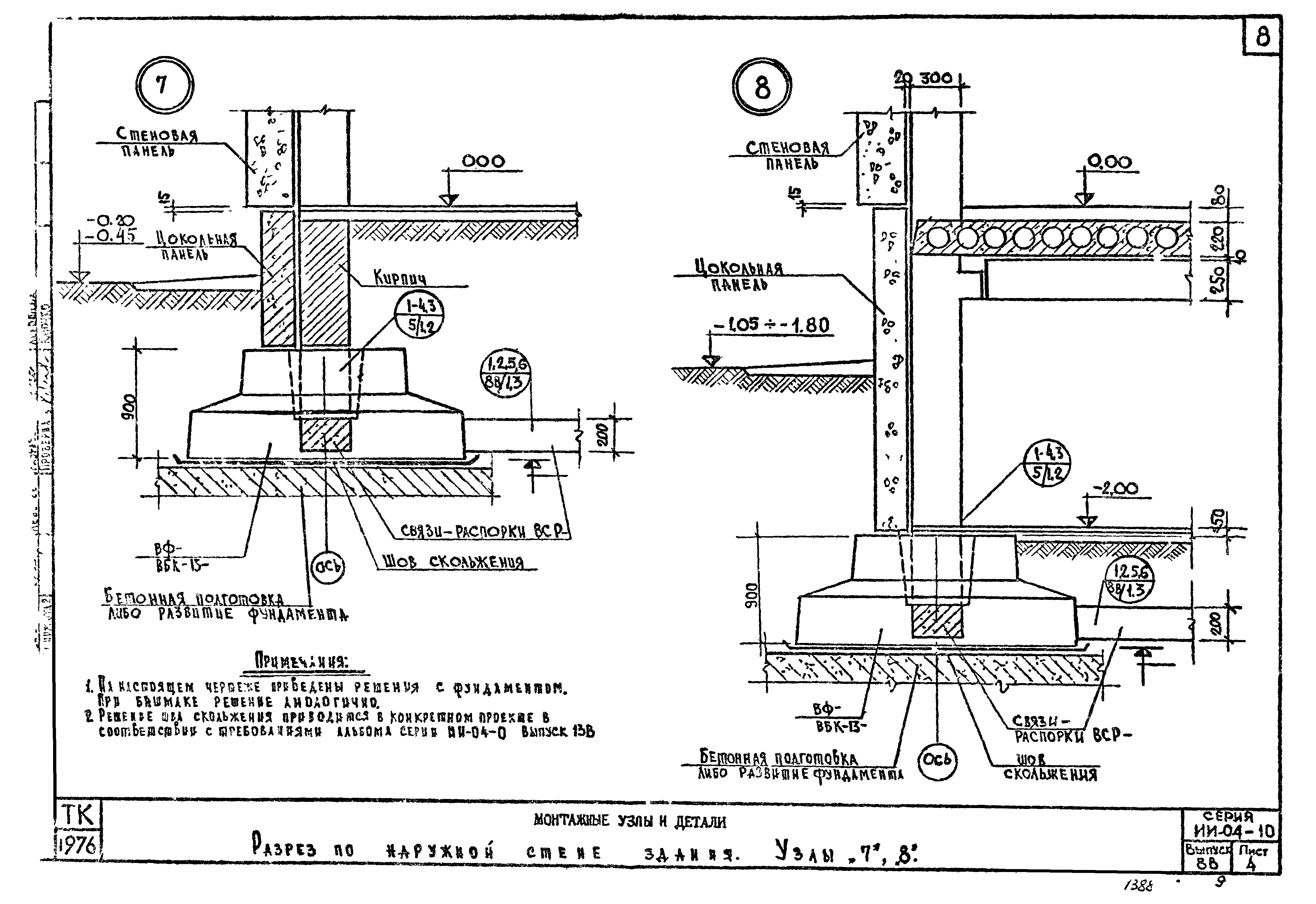
Типовые узлы соединения ростверка со сваей
Типовые узлы соединения ростверка со сваей
После монтажа винтовых свай верхнюю часть фундамента необходимо связать между собой в единую жесткую конструкцию, то есть сделать обвязку – РОСТВЕРК, обвязка также равномерно распределит вес возводимого сооружения на основание.
Ростверк может быть из дерева, метала, бетона.
Ростверк из бруса 200х200
Обвязочный брус (ростверк) притягивается жестко к оголовку сваи через шпильку с диаметром 1012мм. Для того чтобы предотвратить гниение ростверка, брус необходимо обработать антисептиком, также между брусом и оголовком сваи нужно проложить гидроизоляцию желательно на основе нетканых материалов, так как на пяте контакта сваи с брусом будет образоваться конденсат, что приведёт к тому что начнётся загнивание древесины и коррозия железа. После завершения всех сварочных работ на все сворные соединения необходимо нанести антикоррозийный состав.
Вариант 1
Вариант 2
Ростверк из метала (швеллер)
Использование швеллера для обвязки свай:
При использовании швеллера с размером №24 и более для равномерного распределения нагрузок между швеллером и основанием сваи, необходимо к верхнему основанию сваи наварить дополнительно косынки из метала, это дополнительно укрепит, а также распределит нагрузки на верхнее основание винтовой сваи.
Основные типовые узлы для создания конструктивной жесткости фундамента из винтовых свай.
Такие схемы крепления применяются в случаях когда:
При высоте цоколя от 0,7м-1м. Использование сваи: Однолопастная
При высоте цоколя от 1м-1,5м. Использование сваи: Двухлопастная
Сварочные работы должны проводится согласно СНИП 3.03.01-87 «НЕСУЩИЕ И ОГРАЖДАЮЩИЕ КОНСТРУКЦИИ» СП 105-34-96 «Производство сварочных работ и контроль качества сварных соединений» Используемая марка стали для косынок ( ст 3сп толщина 5мм)
Перед тем как приступить сварочным работам, необходимо хорошо зачистить сваи в местах сварочных работ от антикоррозийного покрытия. После завершения всех сварочных работ на все сворные соединения необходимо нанести антикоррозийный состав.
Вариант 1
Вариант 2
Такие схемы крепления применяются в случаях когда:
При высоте цоколя от 1м-1,5м.
 Использование сваи: Однолопастная
Использование сваи: ОднолопастнаяПри высоте цоколя более 1,5м. Использование сваи: Двухлопастная
Сварочные работы должны проводится согласно СНИП 3.03.01-87 «НЕСУЩИЕ И ОГРАЖДАЮЩИЕ КОНСТРУКЦИИ» СП 105-34-96 «Производство работ и контроль качества сварных соединений» Используемая марка стали для косынок ( ст 3сп толщина 5мм)
Перед тем как приступить сварочным работам, необходимо хорошо зачистить сваи в местах предпологаеммых работ от антикоррозийного покрытия. После завершения всех сварочных работ на все сворные соединения необходимо нанести антикоррозийный состав
Вариант 1
Вариант 2
Не удается найти страницу | Autodesk Knowledge Network
(* {{l10n_strings.REQUIRED_FIELD}})
{{l10n_strings.CREATE_NEW_COLLECTION}}*
{{l10n_strings.ADD_COLLECTION_DESCRIPTION}}
{{l10n_strings.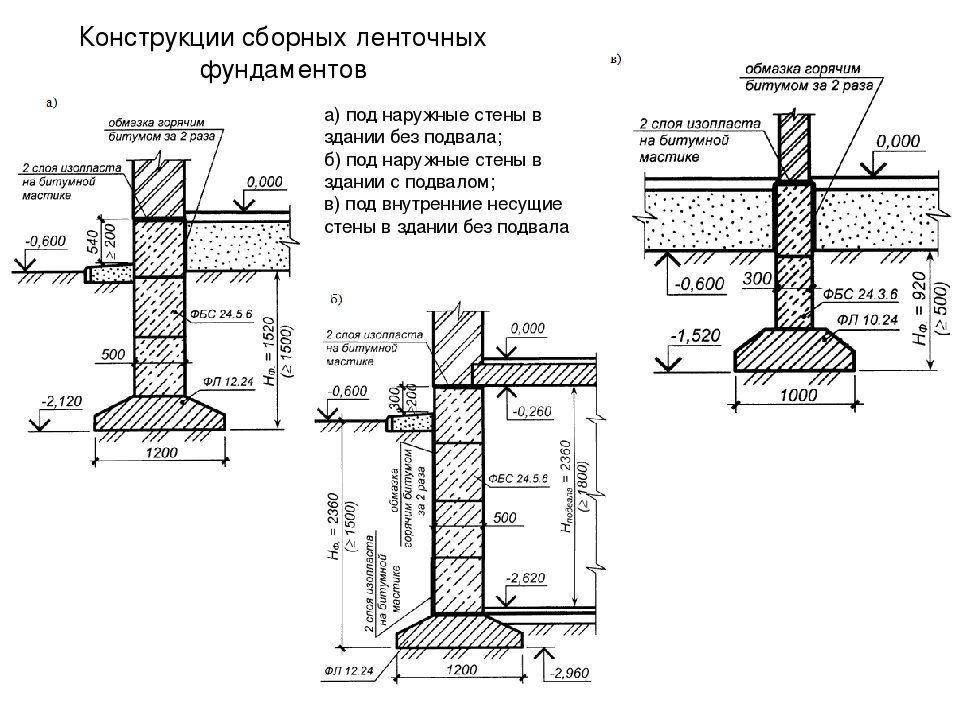
{{l10n_strings.DRAG_TEXT_HELP}}
{{l10n_strings.LANGUAGE}} {{$select.selected.display}}{{article.content_lang.display}}
{{l10n_strings. AUTHOR}}
AUTHOR}}
{{l10n_strings.AUTHOR_TOOLTIP_TEXT}}
{{$select.selected.display}} {{l10n_strings.CREATE_AND_ADD_TO_COLLECTION_MODAL_BUTTON}} {{l10n_strings.CREATE_A_COLLECTION_ERROR}}Подземная гидроизоляция
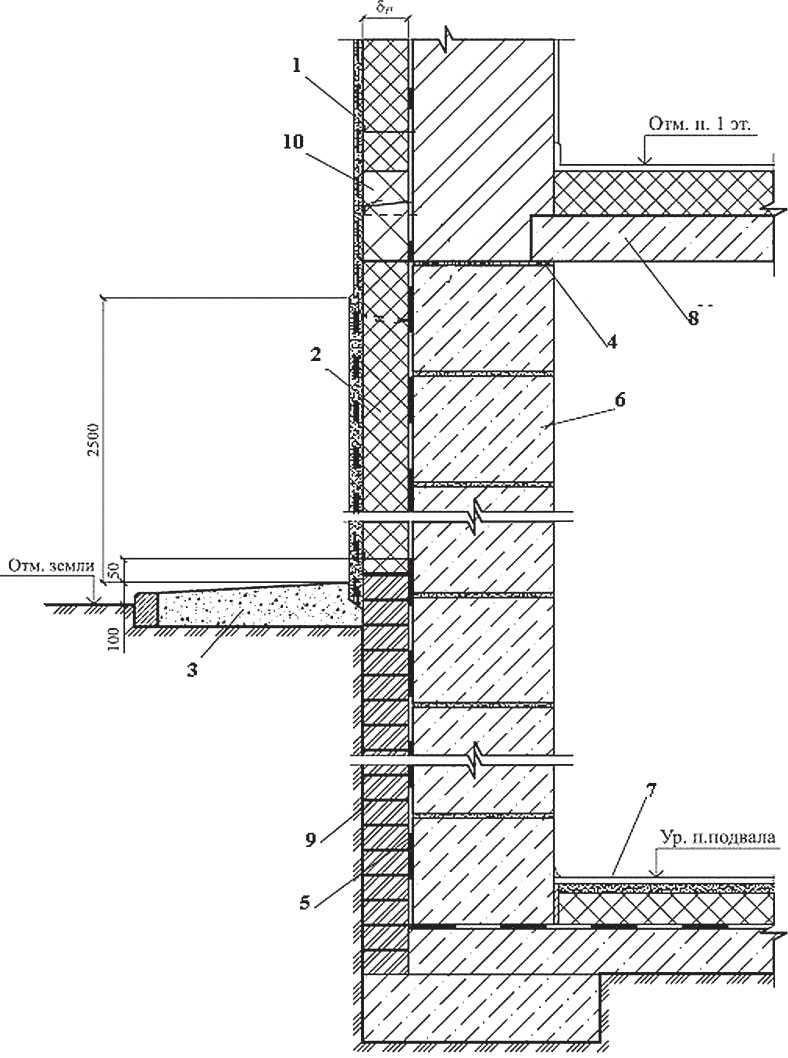
Разработанный техническим отделом Завода КТтрон альбом типовых узлов для проектирования «Гидроизоляция конструкций подземных сооружений материалами «КТтрон» и «КровТрейд» поможет быстро и грамотно запроектировать систему гидроизоляции вашего сооружения.
Подготовленный альбом содержат не только подробно проработанные чертежи узлов и схемы, но и полное описание технологии выполнения работ с типовым сметным расчетом каждого узла.
Скачать альбом «Гидроизоляция конструкций подземных сооружений материалами КТтрон» и «КровТрейд»
Для получения альбома в формате PDF или DWG отправьте запрос по электронной почте [email protected]
Альбом чертежей «Гидроизоляция конструкций подземных сооружений материалами «КТтрон» и «КровТрейд»
Дата актуализации: 17.10.2016
|
Обозначение: |
|
|
Название: |
Гидроизоляция конструкций подземных сооружений материалами «КТтрон» и «КровТрейд» |
|
Статус: |
Действует |
|
Дата введения: |
01. 06.2013 06.2013
|
|
Дата актуализации: |
17.10.2016 |
|
Разработан: |
Технический отдел Завода КТтрон |
Содержание
Раздел 1. Чертежи технических решений по гидроизоляции подземных сооружений при новом строительстве.
Раздел 2. Чертежи технических решений по ремонту гидроизоляции конструкций подземных сооружений
- Узел 18 Схема ремонта трещин
- Узел 19 Схема ремонта и герметизации ввода инженерных коммуникаций
- Узел 20 Схема гидроизоляции по примыканию фундаментная плита-стена
- Схема гидроизоляции монолитного фундамента и наружных кирпичных стен
- Узел 21 Схема гидроизоляции по примыканию фундаментная плита-стен
- Схема гидроизоляции бутового фундамента
- Узел 22 Схема гидроизоляции по примыканию фундаментная плита-стен
- Схема гидроизоляции сборного фундамента с устройством внутреннего дренажа
Раздел 3.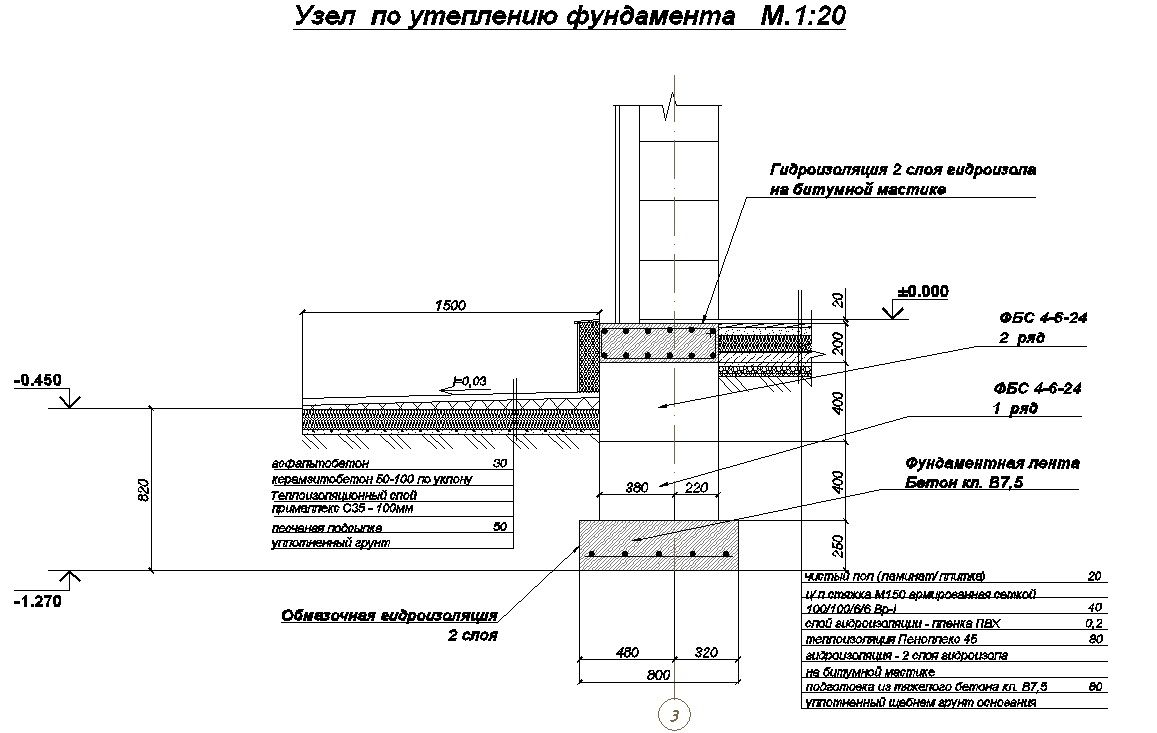 Технология выполнения работ по ремонту трещин
Технология выполнения работ по ремонту трещин
Раздел 4. Локально-сметный расчет технологии работ по гидроизоляции подземных сооружений.
Раздел 5. 3D визуализация чертежей узлов
Скачать альбом «Гидроизоляция конструкций подземных сооружений материалами КТтрон» и «КровТрейд»
Для получения альбома в формате PDF или DWG отправьте запрос по электронной почте [email protected]
Полезные типовые серии – зачем они нужны начинающему проектировщику
Архив рассылки «Непрошеные советы» для начинающих проектировщиков. Выпуск № 27.
Здравствуйте!
С Вами 27 выпуск непрошеных советов. В нем я хочу поговорить о типовых сериях. Очень важно не забывать их ценности, особенно на начальном этапе работы. Даже если серия устарела, в ней можно найти четкую и подробную информацию – до деталей разработанные решения, продуманные и точные узлы. Чтобы не изобретать велосипед, советую начать собирать коллекцию типовых серий, в названиях которые присутствуют «детали» и «узлы». Это самые полезные для начинающего проектировщика шпаргалки.
Чтобы не изобретать велосипед, советую начать собирать коллекцию типовых серий, в названиях которые присутствуют «детали» и «узлы». Это самые полезные для начинающего проектировщика шпаргалки.
Вот, например, ссылки на подборки того, что есть на сайте dwg.ru:
по деталям три страницы;
по узлам четыре страницы.
Чтобы не быть голословной, приведу несколько примеров.
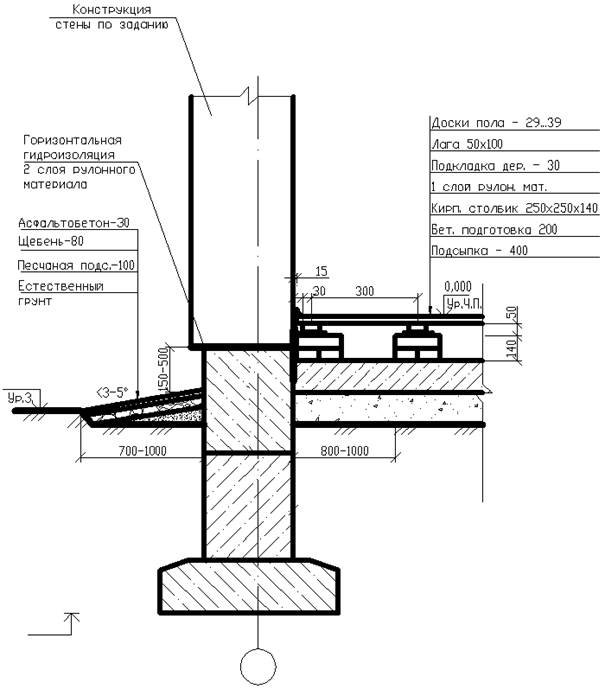
Серия 2.110-1 Детали фундаментов жилых зданий. В этой серии разработаны четкие решения по устройству ленточных фундаментов и стен подвалов – сборных и монолитных. В ней есть и перепады отметок фундамента, и отмостки, и деформационные швы.
Вот сборный фундамент в разрезе: все предельно четко и понятно, как на выставке. И ничего лишнего.
А вот решение по перепаду фундамента. Для многих этот вопрос является проблемой.
Серия 2.140-1 Детали перекрытий жилых зданий. Многие ли знают, что на чертежах перекрытий сборными плитами нужно давать узлы стыковки этих плит между собой и со стенами? А в серии это все доступно показано.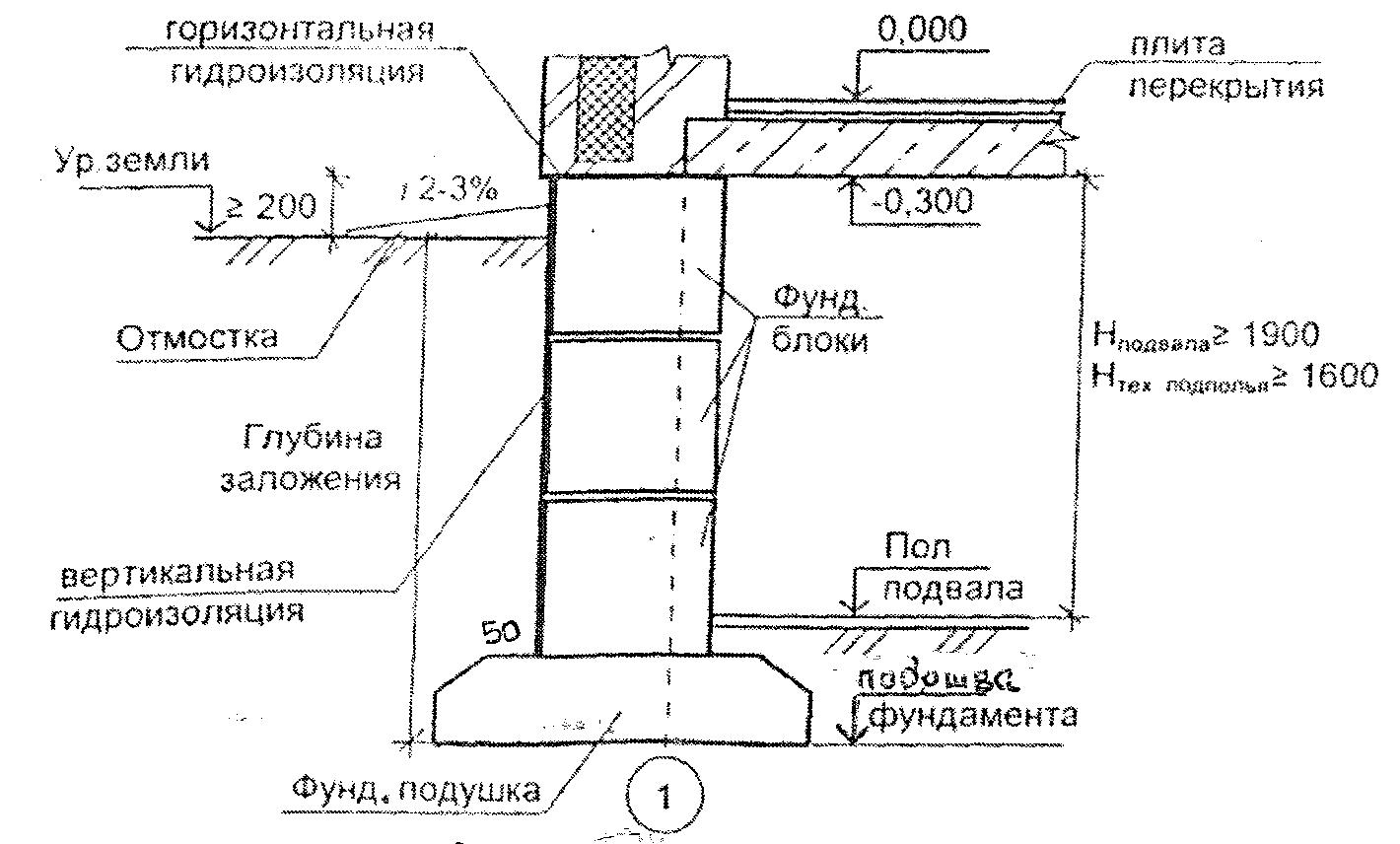
Вот решение по заделке стыков между плитами.
Вот узел опирания плиты на стену – необходимо заказать анкер для плиты и показать его установку. Многие ли это делают?
А вот не менее ценная информация по опиранию пустотных плит на стены с каналами – пустоты в плитах заделываются бетоном, чтобы ароматы и влага никуда не просачивались.
Мой общий совет: скачивайте все типовые серии (особенно с деталями и узлами), которые Вам попадутся, и внимательно их просматривайте – сразу же. Так в голове отложится ссылка на нужную информацию. Вот здесь, кстати, можно скачать перечень типовых серий, по нему легче будет найти для себя нужное.
А вот моя коллекция, делюсь.
И запомните: название серии не всегда отражает ее полное содержание – обычно в ней гораздо больше ценное информации, чем можно себе представить. Но для этого нужно искать, изучать, думать о том, где можно применить.
Успешной Вам творческой работы!
С уважением, Ирина.
— npm
Это порт для стилуса каркаса Zurb Foundation
Вы также можете оформить заказ:
- generator-foundation для быстрого создания настраиваемых тем Foundation.
- fashionista.js, чтобы можно было легко украсить свои приложения express.js с помощью пользовательских тем Foundation.
Foundation — это самая продвинутая в мире адаптивная интерфейсная среда. Вы можете быстро создавать прототипы и создавать сайты или приложения, которые работают на любом устройстве с помощью Foundation, включая конструкции макетов (например, полностью адаптивную сетку), элементы и передовые методы.
Домашняя страница: http://foundation.zurb.com
Документация: http://foundation.zurb.com/docs
Загрузить: http://foundation.zurb.com/download.php
Foundation лицензирован MIT и абсолютно бесплатен для использования. Foundation был бы невозможен без поддержки всей команды ZURB, наших друзей и коллег, которые давали отзывы, и некоторых корифеев, которые проделали тяжелую работу, которой мы воспользовались (спасибо, ребята).
- Базовые исходные файлы и файлы компиляции для SCSS
- Документы
- README
Foundation было создано ZURB, дизайнерской компанией из Кэмпбелла, Калифорния.
Если Foundation сбивает ваши носки так, как мы надеемся, и вы хотите большего, почему бы не проверить наши вакансии?
Большое спасибо всем людям, работающим над Foundation над улучшением базового кода или поддержкой определенных фреймворков. Если вы хотите получить доступ к этому файлу readme, отправьте электронное письмо на адрес [email protected], а если у вас есть вопросы, вы можете присоединиться к группе Google Unofficial Foundation здесь: http://groups.google.com/group/foundation-framework-
WordPress (Версии от 20.03.13)
Joomla
Друпал
Альфред
PyroCMS
Джанго
MODX
.NET
Посредник
Magento
Питон
CodeIgniter
Shopify
Другие реализации
Редакторы
шаблоны
Сетевой генератор
Отображение сетки
Модульные весы
Рубин на рельсах Sass Gems
Генератор Йомена
Настоящим предоставляется бесплатное разрешение любому лицу, получившему копию этого программного обеспечения и связанных файлов документации («Программное обеспечение»), на работу с Программным обеспечением без ограничений, включая, помимо прочего, права на использование, копирование, изменение, объединять, публиковать, распространять, сублицензировать и / или продавать копии Программного обеспечения и разрешать лицам, которым предоставляется Программное обеспечение, делать это при соблюдении следующих условий:
Приведенное выше уведомление об авторских правах и это уведомление о разрешении должны быть включены во все копии или существенные части Программного обеспечения.
ПРОГРАММНОЕ ОБЕСПЕЧЕНИЕ ПРЕДОСТАВЛЯЕТСЯ «КАК ЕСТЬ», БЕЗ КАКИХ-ЛИБО ГАРАНТИЙ, ЯВНЫХ ИЛИ ПОДРАЗУМЕВАЕМЫХ, ВКЛЮЧАЯ, НО НЕ ОГРАНИЧИВАЯСЬ, ГАРАНТИЯМИ КОММЕРЧЕСКОЙ ЦЕННОСТИ, ПРИГОДНОСТИ ДЛЯ ОПРЕДЕЛЕННОЙ ЦЕЛИ И НЕЗАЩИТЫ ОТ ПРАКТИКИ. НИ ПРИ КАКИХ ОБСТОЯТЕЛЬСТВАХ АВТОРЫ ИЛИ ВЛАДЕЛЬЦЫ АВТОРСКИХ ПРАВ НЕ НЕСЕТ ОТВЕТСТВЕННОСТИ ЗА ЛЮБЫЕ ПРЕТЕНЗИИ, УБЫТКИ ИЛИ ДРУГИЕ ОТВЕТСТВЕННОСТЬ, ПРОИЗВОДИМЫЕ В РЕЗУЛЬТАТЕ ДОГОВОРА, ПРАКТИКИ ИЛИ ИНЫМ ОБРАЗОМ, ВОЗНИКАЮЩИМ, ВНУТРИ ИЛИ В СВЯЗИ С ПРОГРАММНЫМ ОБЕСПЕЧЕНИЕМ ИЛИ ИСПОЛЬЗОВАНИЕМ ИЛИ ДРУГИМИ ДЕЛАМИ ПРОГРАММНОЕ ОБЕСПЕЧЕНИЕ.
О нас | Узел.js
В качестве асинхронной среды выполнения JavaScript, управляемой событиями, Node.js предназначен для создания масштабируемые сетевые приложения. В следующем примере «привет, мир» многие соединения могут обрабатываться одновременно. При каждом подключении обратный вызов запущен, но если работы не требуется, Node.js перейдет в спящий режим.
const http = require ('http');
const hostname = '127. 0.0.1';
const порт = 3000;
const server = http.createServer ((req, res) => {
res.statusCode = 200;
res.setHeader ('Content-Type', 'текст / простой');
рез.конец ('Привет, мир');
});
server.listen (порт, имя хоста, () => {
console.log (`Сервер работает по адресу http: // $ {hostname}: $ {port} /`);
});
0.0.1';
const порт = 3000;
const server = http.createServer ((req, res) => {
res.statusCode = 200;
res.setHeader ('Content-Type', 'текст / простой');
рез.конец ('Привет, мир');
});
server.listen (порт, имя хоста, () => {
console.log (`Сервер работает по адресу http: // $ {hostname}: $ {port} /`);
});
Это контрастирует с более распространенной сегодня моделью параллелизма, в которой потоки ОС работают. Сеть на основе потоков относительно неэффективна и очень сложно использовать. Кроме того, пользователи Node.js могут не беспокоиться о тупиковая блокировка процесса, поскольку нет блокировок. Практически нет функции в Node.js напрямую выполняет ввод-вывод, поэтому процесс никогда не блокируется, кроме случаев, когда ввод-вывод выполняется с использованием синхронные методы Node.js стандартная библиотека. Поскольку ничто не блокирует, масштабируемые системы очень разумно разрабатывать на Node.js.
Если кто-то из этого языка незнаком, есть полная статья на
Блокирование и неблокирование.
Node.js похож по дизайну на такие системы, как Ruby, и находится под их влиянием.
Event Machine и Python Twisted. Node.js немного использует модель событий
дальше. Он представляет цикл событий как конструкцию среды выполнения, а не как библиотеку. В других системах
всегда есть блокирующий вызов для запуска цикла событий.Обычно поведение определяется через обратные вызовы в начале скрипта, и
в конце сервер запускается через блокирующий вызов, например EventMachine :: run () .
В Node.js нет такого вызова цикла события. Node.js просто входит в цикл обработки событий после выполнения входного скрипта. Node.js
выходит из цикла обработки событий, когда больше нет обратных вызовов для выполнения. Это поведение
похож на JavaScript в браузере — цикл событий скрыт от пользователя.
HTTP — это первоклассный гражданин в Node.js, разработанный с потоковой передачей и низким
задержка в уме. Это делает Node.js подходящим для создания сети.
библиотека или фреймворк.
Node.js без потоков не означает, что вы не можете
преимущество нескольких ядер в вашей среде. Дочерние процессы могут быть порождены
используя наш child_process.fork () API, и разработаны так, чтобы их было легко
общаться с. На основе того же интерфейса построен модуль cluster ,
который позволяет вам разделять сокеты между процессами, чтобы включить балансировку нагрузки
по вашим ядрам.
Фонд Node.js стал партнером Linux Foundation по новой программе сертификации Node.js
Запуск Node.js Foundation Независимая от поставщика программа сертификации для самой быстрорастущей платформы
САН-ФРАНЦИСКО, 26 января 2017 г. — Фонд Node.js, возглавляемый сообществом и отраслевой консорциум для продвижения разработки платформы Node.js, объявила о разработке первой программы сертификации Node.js направлена на установление базовой компетенции в Node.js.
Node.js — один из самых популярных языков программирования с более чем 4,5 миллионами
активных пользователей в месяц. Хотя Node.js становится все более распространенным
предприятия любого размера сегодня, организации, которые стремятся расширить свои
использование Node.js часто затрудняет переподготовку Java-разработчиков и прием на работу
новый талант.
Хотя Node.js становится все более распространенным
предприятия любого размера сегодня, организации, которые стремятся расширить свои
использование Node.js часто затрудняет переподготовку Java-разработчиков и прием на работу
новый талант.
«Фонд Node.js с помощью невероятных членов сообщества и ядра экспертов, создает комплексную программу сертификации, которая расширяет воронка опытных Node.js имеется опыт. Будь то работа на предприятии среды или в качестве индивидуальных консультантов, тех, кто получил сертификат Node.js Разработчики будут иметь все возможности, чтобы приступить к работе в качестве Node.js. разработчик, обладающий навыками, которые пользуются большим спросом », — сказала Трейси Хиндс, менеджер образовательного сообщества Node.js Foundation.
Программа сертифицированных разработчиков Node.js, которая разрабатывается с участием
Ожидается, что ведущие эксперты и участники Node.js будут доступны во втором квартале 2017 года.Программа предоставит основу для общей компетенции Node. js,
помогая предприятиям быстро находить квалифицированных инженеров Node.js, в то время как
предоставление разработчикам, подрядчикам и консультантам возможности дифференцировать
себя на рынке.
js,
помогая предприятиям быстро находить квалифицированных инженеров Node.js, в то время как
предоставление разработчикам, подрядчикам и консультантам возможности дифференцировать
себя на рынке.
Node.js Foundation тесно сотрудничает с Linux Foundation над созданием проекта и процесс администрирования программы. Linux Foundation предлагает нейтральный дом для проведения программ обучения и сертификации, благодаря его тесное участие в сообществе разработчиков ПО с открытым исходным кодом.Он предлагает несколько открытых онлайн курсы (MOOC), включая Введение в Linux, Введение в DevOps: Преобразование и улучшение операций; Разработка Приложения для Linux; Kubernetes Основы; среди многих других.
Ideal Node.js Certified Developer кандидаты — первые разработчики среднего уровня. кто уже может профессионально работать с JavaScript на платформе Node.js. Стоимость самостоятельного онлайн-экзамена остается прежней. быть определенным.
В настоящее время Фонд Node.js работает с сообществом, чтобы определить конкретные вопросы, которые будут заданы на экзамене. К
внести свой вклад в написание элементов разработки для сертификации Node.js Foundation
Workshop Sessions, заполните эту заявку.
К
внести свой вклад в написание элементов разработки для сертификации Node.js Foundation
Workshop Sessions, заполните эту заявку.
тем экзамена будут опубликованы в открытом доступе, как и ресурсы, которые помогут подготовиться к экзамену. сертификация, позволяющая другим использовать исходные материалы для собственных Обучение Node.js.
Основы Node.js и JS хотят объединить — TechCrunch
В настоящее время существует два основных фонда с открытым исходным кодом, которые сосредоточены на JavaScript: JS Foundation, основанная в 2016 году, и Node.js Foundation, который был запущен в 2015 году. Миссия JS Foundation состоит в том, чтобы управлять экосистемой вокруг языка, в то время как Node.js, очевидно, фокусируется на технологии Node.js для использования JavaScript на стороне сервера с помощью движка Google V8 и развития этой экосистемы. . Теперь эти два фонда хотят слиться.
Это еще не сделано, и две организации планируют получить отзывы от своих сообществ, начиная с личных вопросов и ответов на предстоящей конференции Node + JS Interactive, а также онлайн.
«Объединение сил не повлияет на техническую независимость или автономию Node.js или любого из 28 проектов JS Foundation, таких как Appium, ESLint или jQuery», — пишут обе организации в сегодняшнем заявлении. «JavaScript — это универсальный язык программирования, который вышел далеко за рамки своей роли в качестве основы Интернета, войдя в новые среды, такие как IoT, собственные приложения, DevOps и протоколы. По мере того, как экосистема продолжает развиваться — переходя от браузеров к серверам, от настольных приложений к встроенным устройствам — усиление сотрудничества в экосистеме JavaScript становится как никогда важным для поддержания непрерывного и здорового роста.”
И действительно, это расширение сотрудничества в экосистеме, по-видимому, лежит в основе этого шага. «Руководители Фонда и ключевые технические заинтересованные стороны считают, что более тесное объединение сообществ расширит сферу действия текущих Основ и обеспечит большую поддержку Node.js и более широкого круга проектов JavaScript», — отметил Майк Долан, вице-президент по стратегическим программам в Linux Foundation в сегодняшнем объявлении.
Конечными целями слияния являются «повышение операционной эффективности», расширение сотрудничества и расширение сотрудничества в экосистемах JavaScript, а также создание единой организации, которая может стать домом для любого проекта JavaScript.
Если это слияние произойдет, то оно, несомненно, поможет обеим группам, которые уже находятся под эгидой Linux Foundation, что должно сделать переход относительно легким при условии согласия сообщества. Есть также некоторое совпадение между членами двух групп, хотя, вероятно, меньше, чем вы ожидаете. В то время как IBM является платиновым членом обоих, например, Google является платиновым членом фонда Node.JS, но не спонсирует JS Foundation. Точно так же Samsung является спонсором высшего уровня JS Foundation, но его нигде нет на Node.Сайт JS Foundations. Тогда, вероятно, неудивительно, что одной из заявленных целей слияния также является «оптимизация взаимодействия с участниками».
Количественная протеомика и одноядерная транскриптомика синусового узла проясняет основы кардиостимуляции
Сбор тканей
Тридцать самцов мышей C57BL / 6J в возрасте 12–14 недель и массой тела 25–35 г были последовательно умерщвлены цервикальным путем. дислокации в соответствии с Законом Соединенного Королевства о животных (научные процедуры) 1986 года и Директивой 2010/63 / ЕС Европейского парламента.Уход и использование животных осуществлялись в соответствии со стандартами и практиками, утвержденными органом по защите животных и этике Манчестерского университета, и в соответствии с Законом о животных (научные процедуры) 1986 года. Сердца были быстро удалены, а правое предсердие вместе с целым синусового узла (рис. 1b) иссекали в холодном фосфатно-солевом буфере (Sigma-Aldrich, UK). Для синусового узла иссекали полупрозрачную межполостную область между концевой кристой и межпредсердной перегородкой (~ 1 на 1 мм).В отдельной когорте сердец срезы тканей через полупрозрачную межполостную область были помечены иммуномечением для активированных гиперполяризацией циклических нуклеотид-зависимых каналов (HCN4) и белков коннексина 43 (Cx43) с использованием протоколов, описанных ранее 76 и описанных ниже в разделе иммуногистохимии.
дислокации в соответствии с Законом Соединенного Королевства о животных (научные процедуры) 1986 года и Директивой 2010/63 / ЕС Европейского парламента.Уход и использование животных осуществлялись в соответствии со стандартами и практиками, утвержденными органом по защите животных и этике Манчестерского университета, и в соответствии с Законом о животных (научные процедуры) 1986 года. Сердца были быстро удалены, а правое предсердие вместе с целым синусового узла (рис. 1b) иссекали в холодном фосфатно-солевом буфере (Sigma-Aldrich, UK). Для синусового узла иссекали полупрозрачную межполостную область между концевой кристой и межпредсердной перегородкой (~ 1 на 1 мм).В отдельной когорте сердец срезы тканей через полупрозрачную межполостную область были помечены иммуномечением для активированных гиперполяризацией циклических нуклеотид-зависимых каналов (HCN4) и белков коннексина 43 (Cx43) с использованием протоколов, описанных ранее 76 и описанных ниже в разделе иммуногистохимии. Полупрозрачная межполостная область помечена положительной для HCN4 и отрицательной для Cx43, что типично для синусового узла (рис. 1b). Для правого предсердия была рассечена часть ушка правого предсердия (~ 1 на 1 мм).Образцы тканей помещали в криопробирки и быстро замораживали в жидком азоте. Образцы от десяти животных объединяли в каждую пробирку, получая по три пробирки для синусового узла и предсердной мышцы. Объединение образцов было вызвано небольшим размером синусового узла мыши и необходимостью сбора достаточного количества белка для обнаружения с помощью масс-спектрометрии. Образцы тканей отправляли на сухом льду и хранили при -80 ° C до использования. Последующие эксперименты и анализ данных проводились вслепую.
Полупрозрачная межполостная область помечена положительной для HCN4 и отрицательной для Cx43, что типично для синусового узла (рис. 1b). Для правого предсердия была рассечена часть ушка правого предсердия (~ 1 на 1 мм).Образцы тканей помещали в криопробирки и быстро замораживали в жидком азоте. Образцы от десяти животных объединяли в каждую пробирку, получая по три пробирки для синусового узла и предсердной мышцы. Объединение образцов было вызвано небольшим размером синусового узла мыши и необходимостью сбора достаточного количества белка для обнаружения с помощью масс-спектрометрии. Образцы тканей отправляли на сухом льду и хранили при -80 ° C до использования. Последующие эксперименты и анализ данных проводились вслепую.
Гомогенизация тканей
Биоптаты замороженных тканей гомогенизировали на гомогенизаторе Precellys24 (Bertin Technologies, Франция) в буфере тритона x-100 на основе Триса (50 мМ Трис-HCl, pH 8.5, 5 мМ ЭДТА, 150 мМ NaCl, 10 мМ KCl, 1% Тритон X-100, 5 мМ NaF, 5 мМ бета-глицерофосфат, 1 мМ Na-ортованадат, содержащий 1x полный ингибитор протеазы Roche) с керамическими шариками (2,8 и Шарики из оксида циркония 1,4 мм, Precellys). Гомогенаты инкубировали в течение 2 ч при 4 ° C (20 об / мин), а затем переносили в охлажденные пробирки объемом 1,5 мл. Лизаты тканей обрабатывали ультразвуком в течение пяти циклов (30 секунд включения / 30 секунд при низкой амплитуде) в ультразвуковом аппарате Bioruptor (Diagenode, США), очищали центрифугированием (15000 × г , 20 минут, 4 ° C) и собирали растворимые фракции. .Для удаления поверхностно-активных веществ перед измерением ЖХ-МС / МС белок осаждали добавлением трихлоруксусной кислоты (ТХК) до конечной концентрации 10%. Образцы инкубировали в течение 30 минут на льду и осторожно центрифугировали (200 × g , 20 с) для сбора белка. Супернатанты отбрасывали, и белок ресуспендировали в буфере гуанидин-HCl (Gnd-HCl; 6MGnd-HCl, 50 мМ Tris-HCl pH 8,5, 5 мМ NaF, 5 мМ бета-глицерофосфат, 1 мМ Na-ортованадат, содержащий 1x полный ингибитор протеазы Roche. ). Дисульфидные мостики восстанавливали и цистеиновые фрагменты алкилировали добавлением 5 мМ трис (2-карбоксиэтил) фосфина (TCEP) и 10 мМ хлорацетамида (CAA) и инкубацией в темноте при комнатной температуре в течение 15 минут.
Гомогенаты инкубировали в течение 2 ч при 4 ° C (20 об / мин), а затем переносили в охлажденные пробирки объемом 1,5 мл. Лизаты тканей обрабатывали ультразвуком в течение пяти циклов (30 секунд включения / 30 секунд при низкой амплитуде) в ультразвуковом аппарате Bioruptor (Diagenode, США), очищали центрифугированием (15000 × г , 20 минут, 4 ° C) и собирали растворимые фракции. .Для удаления поверхностно-активных веществ перед измерением ЖХ-МС / МС белок осаждали добавлением трихлоруксусной кислоты (ТХК) до конечной концентрации 10%. Образцы инкубировали в течение 30 минут на льду и осторожно центрифугировали (200 × g , 20 с) для сбора белка. Супернатанты отбрасывали, и белок ресуспендировали в буфере гуанидин-HCl (Gnd-HCl; 6MGnd-HCl, 50 мМ Tris-HCl pH 8,5, 5 мМ NaF, 5 мМ бета-глицерофосфат, 1 мМ Na-ортованадат, содержащий 1x полный ингибитор протеазы Roche. ). Дисульфидные мостики восстанавливали и цистеиновые фрагменты алкилировали добавлением 5 мМ трис (2-карбоксиэтил) фосфина (TCEP) и 10 мМ хлорацетамида (CAA) и инкубацией в темноте при комнатной температуре в течение 15 минут.
Пептидный препарат
Из каждого образца до 1 мг белка переваривали в растворе путем добавления эндопротеиназы Lys-C (Trichem ApS, Дания; соотношение фермент: белок 1: 100) в течение 1 ч при 30 ° C, 750 об / мин в темноте с последующим разбавлением (1:12 50 мМ Трис-HCl, pH 8) и расщеплением трипсином в течение ночи (16 ч) при 37 ° C, 750 об / мин (Life Technologies, США, соотношение фермент: белок 1: 100 ). Реакции гасили добавлением трифторуксусной кислоты (TFA, конечная концентрация 1%) и образцы центрифугировали (14000 × г, , 10 мин) до остатков осадка.Растворимые фракции обессоливали и концентрировали на колонках C18 SepPak (Waters, США). Пептиды элюировали 40% ацетонитрилом (ACN), затем 60% ACN, а затем органические растворители выпаривали центрифугированием в вакууме.
Автономное фракционирование образцов пептида с высоким pH
Из каждого образца 100 мкг пептида (в объеме инъекции 10 мкл) фракционировали с помощью микропоточной жидкостной хроматографии сверхвысокого давления с обращенной фазой (UPLC) на системе Dionex UltiMate 3000 UPLC (Thermo Scientific, США), оборудованную колонкой ACQUITY UPLC CSH C18 (130Å, 1. 7 мкм, 1 мм x 150 мм) при скорости потока 30 мкл / мин 6,7 . Использовалась следующая программа градиентного элюирования 85 мин с использованием бинарного насоса, подключенного к растворителю A (5 мМ бикарбонат аммония (ABC), pH 8) и B (100% ACN): 0–50 мин: 4,5–22,5% B, 50– 55 мин: 22,5–63% B, 55–60 мин: 63% B изократический, 60–62 мин: 63–81% B, 62–70 мин: 81% B изократический, 70–75 мин: 81–4,5% B с последующим повторным уравновешиванием колонки при 4,5% B в течение 10 мин. Отток через 4–60 мин собирали с интервалами в 1 мин в 12 объединенных фракций в автосэмплере.Фракции подкисляли добавлением 5 мкл 5% муравьиной кислоты (FA), чтобы избежать осаждения ABC, и объем фракции уменьшали центрифугированием в вакууме. Если не указано иное, химические вещества и реагенты были приобретены у Sigma-Aldrich, США. Растворители для хроматографии были приобретены в VWR, США.
7 мкм, 1 мм x 150 мм) при скорости потока 30 мкл / мин 6,7 . Использовалась следующая программа градиентного элюирования 85 мин с использованием бинарного насоса, подключенного к растворителю A (5 мМ бикарбонат аммония (ABC), pH 8) и B (100% ACN): 0–50 мин: 4,5–22,5% B, 50– 55 мин: 22,5–63% B, 55–60 мин: 63% B изократический, 60–62 мин: 63–81% B, 62–70 мин: 81% B изократический, 70–75 мин: 81–4,5% B с последующим повторным уравновешиванием колонки при 4,5% B в течение 10 мин. Отток через 4–60 мин собирали с интервалами в 1 мин в 12 объединенных фракций в автосэмплере.Фракции подкисляли добавлением 5 мкл 5% муравьиной кислоты (FA), чтобы избежать осаждения ABC, и объем фракции уменьшали центрифугированием в вакууме. Если не указано иное, химические вещества и реагенты были приобретены у Sigma-Aldrich, США. Растворители для хроматографии были приобретены в VWR, США.
Измерения ЖХ-МС / МС
Образцы фракционированных пептидов анализировали с помощью обращенно-фазовой жидкостной хроматографии в режиме онлайн, подключенной к тандемному масс-спектрометру Q-Exactive HF quadrupole Orbitrap (Thermo Electron, Бремен, Германия). Образцы пептидов доводили до концентрации 0,2 мкг / мкл (разбавляли в 5% ACN, 0,1% TFA) в 96-луночных микротитрационных планшетах и автоматически отбирали (объем инъекции 5 мкл) в систему nanoflow Easy-nLC (Proxeon Biosystems, Odense, Дания. ). Образцы пептидов разделяли на эмиттерных колонках из плавленого кварца 15 см (внутренний диаметр 75 мкм), вытягивали и набивали на месте обращенно-фазовой смолой ReproSil-Pur C18-AQ 1,9 мкм (Dr. Maisch GmbH, Аммербух-Энтринген, Германия) в многоступенчатый линейный градиент в течение 1 часа (константа муравьиной кислоты 0,1%; 2-25% ACN за 45 минут, 25-45% ACN за 8 минут, 45-80% ACN за 3 минуты) с последующим повторным уравновешиванием короткой колонки ( 80–5% ACN за 5 минут, 5% ACN за 2 минуты).Выходящий из колонки поток подвергали прямой ионизации в источнике ионизации с наноэлектрораспылением, работающем в режиме положительной ионизации, и подвергали электрораспылению в масс-спектрометр. Спектры полного МС (375–1500 m / z) были получены после накопления 3 000 000 ионов в Orbitrap (максимальное время заполнения 25 мс) при разрешении 120 000.
Образцы пептидов доводили до концентрации 0,2 мкг / мкл (разбавляли в 5% ACN, 0,1% TFA) в 96-луночных микротитрационных планшетах и автоматически отбирали (объем инъекции 5 мкл) в систему nanoflow Easy-nLC (Proxeon Biosystems, Odense, Дания. ). Образцы пептидов разделяли на эмиттерных колонках из плавленого кварца 15 см (внутренний диаметр 75 мкм), вытягивали и набивали на месте обращенно-фазовой смолой ReproSil-Pur C18-AQ 1,9 мкм (Dr. Maisch GmbH, Аммербух-Энтринген, Германия) в многоступенчатый линейный градиент в течение 1 часа (константа муравьиной кислоты 0,1%; 2-25% ACN за 45 минут, 25-45% ACN за 8 минут, 45-80% ACN за 3 минуты) с последующим повторным уравновешиванием короткой колонки ( 80–5% ACN за 5 минут, 5% ACN за 2 минуты).Выходящий из колонки поток подвергали прямой ионизации в источнике ионизации с наноэлектрораспылением, работающем в режиме положительной ионизации, и подвергали электрораспылению в масс-спектрометр. Спектры полного МС (375–1500 m / z) были получены после накопления 3 000 000 ионов в Orbitrap (максимальное время заполнения 25 мс) при разрешении 120 000. Был использован зависимый от данных метод Top12, позволяющий проводить глубокие измерения протеома 8 , последовательно выделяя наиболее интенсивные ионы-предшественники (до 12 за полное сканирование) для более высокой энергии столкновительной диссоциации (HCD) в ячейке столкновения октополя.МС / МС спектры фрагментных ионов были записаны с разрешением 30 000 после накопления 100 000 ионов в Orbitrap (максимальное время заполнения 45 мс).
Был использован зависимый от данных метод Top12, позволяющий проводить глубокие измерения протеома 8 , последовательно выделяя наиболее интенсивные ионы-предшественники (до 12 за полное сканирование) для более высокой энергии столкновительной диссоциации (HCD) в ячейке столкновения октополя.МС / МС спектры фрагментных ионов были записаны с разрешением 30 000 после накопления 100 000 ионов в Orbitrap (максимальное время заполнения 45 мс).
Обработка необработанных данных
Необработанные данные МС были обработаны с использованием программного обеспечения MaxQuant v1.5.3.30 77 и белков, идентифицированных с помощью встроенной поисковой системы Andromeda путем поиска спектров МС / МС по триптическому гидролизу in silico базы данных содержащий все проверенные записи о белках SwissProt (загружено 6.8.2016). Спектры МС / МС исследовали с карбамидометлицистеином в качестве фиксированной модификации, а также с окислением (M), ацетилированием N-концов белка и Gln-> пиро-Glu в качестве вариабельных модификаций. Допускалось максимум два пропущенных расщепления и шесть вариабельных модификаций. Минимальная длина пептида была установлена равной семи аминокислотам (по умолчанию), а минимальная оценка Андромеды, необходимая для модифицированных пептидов, составляла 25, с минимальной дельта-оценкой 6 (по умолчанию). Допуск первого поиска составлял 20 ppm (по умолчанию), а основной допуск поиска составлял 4,5 ppm (по умолчанию), что требовало строгой специфичности триптических пептидов. Из-за схожести образцов опция сопоставления между запусками была включена с параметрами по умолчанию.Порог ложного обнаружения был установлен на 1% на уровне пептидов, белков и ложных сайтов (по умолчанию), что позволяло проходить только высококачественные идентификации. Все пептиды использовали для количественного определения белка, а количественное определение без метки (LFQ) было дополнительно выполнено в MaxQuant с включенной опцией быстрого LFQ. Результаты идентификации белков были дополнительно обработаны с использованием программного пакета Perseus 78 версии 1.
Допускалось максимум два пропущенных расщепления и шесть вариабельных модификаций. Минимальная длина пептида была установлена равной семи аминокислотам (по умолчанию), а минимальная оценка Андромеды, необходимая для модифицированных пептидов, составляла 25, с минимальной дельта-оценкой 6 (по умолчанию). Допуск первого поиска составлял 20 ppm (по умолчанию), а основной допуск поиска составлял 4,5 ppm (по умолчанию), что требовало строгой специфичности триптических пептидов. Из-за схожести образцов опция сопоставления между запусками была включена с параметрами по умолчанию.Порог ложного обнаружения был установлен на 1% на уровне пептидов, белков и ложных сайтов (по умолчанию), что позволяло проходить только высококачественные идентификации. Все пептиды использовали для количественного определения белка, а количественное определение без метки (LFQ) было дополнительно выполнено в MaxQuant с включенной опцией быстрого LFQ. Результаты идентификации белков были дополнительно обработаны с использованием программного пакета Perseus 78 версии 1. 5.5.3 и среды программирования R (R v3.3.3 в RStudio v1.0.136). Квантильная нормализация исходных интенсивностей была выполнена на основе пакета Bioconductor R LIMMA 79 .Для количественного анализа белковые группы, представляющие изоформы одного и того же канонического белка, были удалены из данных, так что была сохранена только самая распространенная изоформа белка.
5.5.3 и среды программирования R (R v3.3.3 в RStudio v1.0.136). Квантильная нормализация исходных интенсивностей была выполнена на основе пакета Bioconductor R LIMMA 79 .Для количественного анализа белковые группы, представляющие изоформы одного и того же канонического белка, были удалены из данных, так что была сохранена только самая распространенная изоформа белка.
Количественный анализ
Общий анализ значимых различий между образцами синусового узла (SN) и правого предсердия (RA) был проведен в Perseus на значениях LFQ технической реплики A из-за лучшего общего качества данных этой реплики. Обратные идентификации и определения примесей были удалены из данных, а информация об изоформах была объединена в каноническую группу белков (т.е.е., если было идентифицировано более одной изоформы, рассматривалась только изоформа с наивысшей интенсивностью). Анализ главных компонентов (PCA) проводился на 100% достоверных значениях. Для всех других количественных анализов данные были отфильтрованы так, чтобы были сохранены только белки с по крайней мере тремя действительными значениями по крайней мере в одной из областей сердца (SN или RA).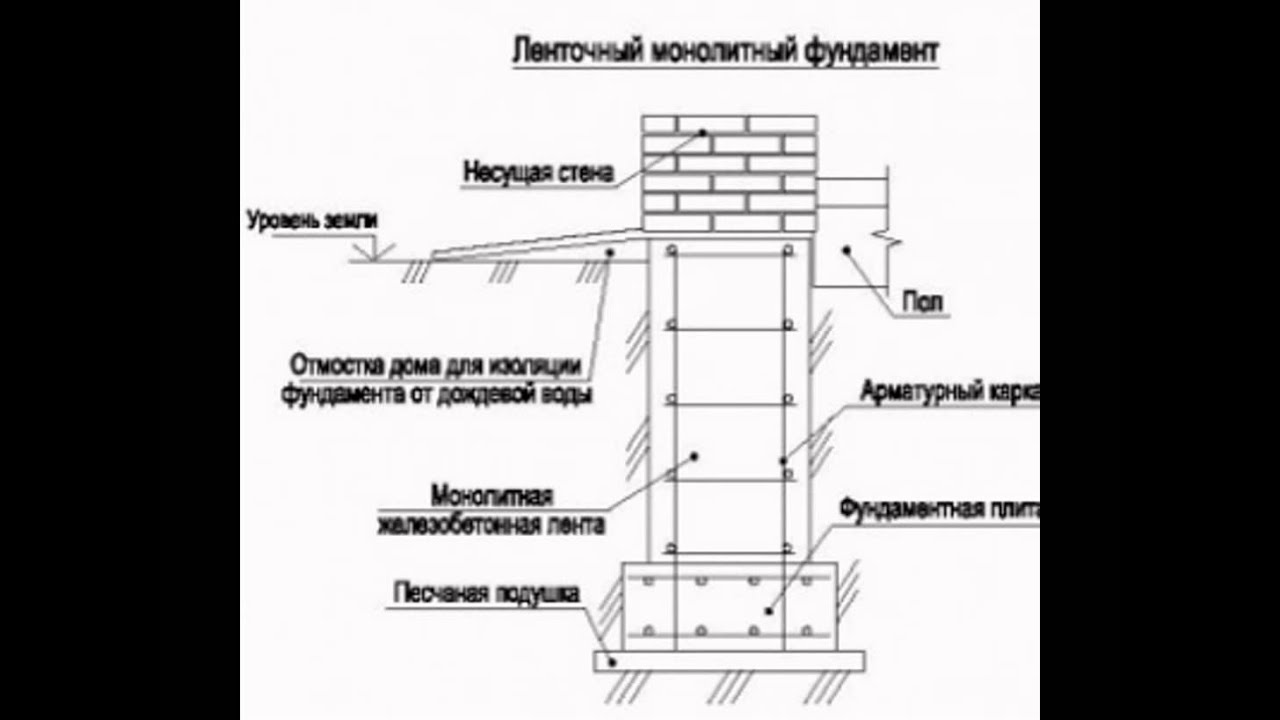
Анализ графика вулкана
Для глобального анализа количественных различий (график вулкана) пропущенные значения были рассчитаны отдельно для каждого столбца из нижнего конца оригинала.В частности, вменение отсутствующих значений было выполнено в программном обеспечении Perseus со стандартными настройками: значения были взяты из распределения с шириной 0,3 и сдвигом вниз 1,8 по сравнению с исходным распределением выборки. Вменение производилось отдельно для каждого образца. 5523 белка, оставшиеся после фильтрации, дали 33 138 теоретических значений (6 × 5523), из которых 32 433 были действительными (измеренными), а остальные 819 были условными (~ 2,5% от общего количества). Максимальное количество вмененных значений для одной выборки составляло 194 значения (SN1), за которыми следовали SN3 (190), SN2 (178), RA1 (94), RA2 (82) и RA3 (81 значение).В последующий анализ были включены только белки, идентифицированные по крайней мере с двумя пептидами. Анализ графика вулкана Персей проводился с применением двустороннего теста Стьюдента t для проверки значимой разницы в содержании белка между SN и RA. Отсечка частоты ложных обнаружений (FDR) на основе перестановок была определена с 750 рандомизацией и S 0 = 0,1 (по умолчанию). Из 5523 белков, оставшихся после фильтрации, 587 считались значимыми при 5% FDR. Среди белков с вмененными значениями мы впоследствии вручную проверили значимые белки, чтобы убедиться, что значимость никогда не была основана на вменении из-за плохой количественной оценки.С этой целью мы исследовали необработанные интенсивности всех значимых белков, которые имели только 2, 1 или нулевые значения LFQ в SN или RA. Если мы записали необработанные интенсивности, но они не были количественно определены в LFQ, мы дважды проверили, была ли значимость только из-за потери точек данных во время количественной оценки (и последующего вменения), т. Е. Если соотношение исходных интенсивностей интенсивностей LFQ между SN и RA сильно отклонялись от соотношения, наблюдаемого после вменения. На этом основании мы решили исключить список из 12 белков из дальнейших анализов, которые с большой вероятностью считаются ложноположительными из-за сильно завышенной разницы кратных изменений (SN / RA) по сравнению с необработанными интенсивностями: Cyb5d2, Fxyd1, Gfra2, Ca4, Ntm, Mup3, Sema7a, Ddx18, Fgd4, Gas2l1, Ptpn18 и Cend1.
Отсечка частоты ложных обнаружений (FDR) на основе перестановок была определена с 750 рандомизацией и S 0 = 0,1 (по умолчанию). Из 5523 белков, оставшихся после фильтрации, 587 считались значимыми при 5% FDR. Среди белков с вмененными значениями мы впоследствии вручную проверили значимые белки, чтобы убедиться, что значимость никогда не была основана на вменении из-за плохой количественной оценки.С этой целью мы исследовали необработанные интенсивности всех значимых белков, которые имели только 2, 1 или нулевые значения LFQ в SN или RA. Если мы записали необработанные интенсивности, но они не были количественно определены в LFQ, мы дважды проверили, была ли значимость только из-за потери точек данных во время количественной оценки (и последующего вменения), т. Е. Если соотношение исходных интенсивностей интенсивностей LFQ между SN и RA сильно отклонялись от соотношения, наблюдаемого после вменения. На этом основании мы решили исключить список из 12 белков из дальнейших анализов, которые с большой вероятностью считаются ложноположительными из-за сильно завышенной разницы кратных изменений (SN / RA) по сравнению с необработанными интенсивностями: Cyb5d2, Fxyd1, Gfra2, Ca4, Ntm, Mup3, Sema7a, Ddx18, Fgd4, Gas2l1, Ptpn18 и Cend1.
Сетевой анализ
Чтобы исследовать ассоциации между значимыми белками, мы извлекли белковые ассоциации из базы данных STRING 80 для всех значимых белков из анализа графика вулкана. STRING идентифицировал 566/575 значимых белков и вернул 2176 взаимодействий при пороговой достоверности 0,4 (средний уровень достоверности для STRING). Никаких взаимодействий не было возвращено для 91 белка, тогда как 463 сгруппированы в основную сеть, а остальные десять — в более мелкие отдельные сети из 2–3 белков.Мы выполнили кластеризацию MCL в сети в Cytoscape 81 , используя Clustermaker2 82 . Кластеризация проводилась относительно оценки при коэффициенте инфляции 3,0, так что взаимодействия с более высокой оценкой имели больший вес во время кластеризации. Из 109 возвращенных кластеров 24 кластера содержали пять или более членов (всего 228 белков). Анализ обогащения онтологии был выполнен для кластеров с пятью или более членами с использованием приложения STRING (v1. 1.0) в Cytoscape 80,81 .На основе анализа обогащения были выведены общие термины, описывающие основные различия между синусовым узлом и кластерами правого предсердия. Для анализа меньших групп функционально связанных белков мы извлекли белковые ассоциации для белков человека из STRING и наложили данные об экспрессии белков мыши. Уровень достоверности был установлен на средний уровень достоверности (0,4) во всех сетях, кроме сети с потенциалом действия; здесь граница оценки была снижена до 0,3, чтобы включить ассоциацию HCN4-HCN1.
1.0) в Cytoscape 80,81 .На основе анализа обогащения были выведены общие термины, описывающие основные различия между синусовым узлом и кластерами правого предсердия. Для анализа меньших групп функционально связанных белков мы извлекли белковые ассоциации для белков человека из STRING и наложили данные об экспрессии белков мыши. Уровень достоверности был установлен на средний уровень достоверности (0,4) во всех сетях, кроме сети с потенциалом действия; здесь граница оценки была снижена до 0,3, чтобы включить ассоциацию HCN4-HCN1.
Иммуногистохимия
Препараты ткани правого предсердия с интактным синусовым узлом ( n = 2; типичный препарат, показанный на рис. 1b) заключали в соединение OCT (VWR International, UK) и замораживали путем заливки 2-метилбутана (Sigma-Aldrich) , UK) охлаждаемые в жидкости N 2 . Замороженные серийные срезы (толщиной 20 мкм) вырезали перпендикулярно концевой кристе от проксимального до дистального конца препаратов в криостате Leica CM3050 S (Leica Microsystems, Германия) при -18 ° C. Вырезанные срезы помещали на предметные стекла SuperFrost Plus (VWR International, UK) и хранили при -80 ° C до дальнейшего использования. Замороженные серийные срезы фиксировали в 10% формалине, приготовленном в фосфатно-солевом буфере (PBS; Sigma-Aldrich, UK), в течение 30 минут, а затем промывали PBS (Sigma-Aldrich, UK) три раза (10 минут / промывка). Ткани были проницаемы с помощью 0,1% Triton X-100 в PBS (Sigma-Aldrich, UK) в течение 30 минут, трижды промыли PBS (10 минут / промывка) и заблокировали 1% бычьим сывороточным альбумином (BSA; Sigma-Aldrich, UK). ) в PBS в течение ~ 30 мин.BSA удаляли, и срезы тканей инкубировали при 4 ° C в течение ночи с соответствующими первичными антителами, разведенными в 1% BSA в PBS. Все антитела, использованные в этом исследовании, были коммерчески доступны, проверены поставщиками на видовую реактивность и для иммунофлуоресценции и перечислены в дополнительной таблице 1. Были использованы разведения, рекомендованные поставщиком. После инкубации с первичными антителами в течение ночи срез ткани трижды промывали PBS (10 мин / промывка) и инкубировали в течение 2 ч при комнатной температуре с соответствующими вторичными антителами (см.
Вырезанные срезы помещали на предметные стекла SuperFrost Plus (VWR International, UK) и хранили при -80 ° C до дальнейшего использования. Замороженные серийные срезы фиксировали в 10% формалине, приготовленном в фосфатно-солевом буфере (PBS; Sigma-Aldrich, UK), в течение 30 минут, а затем промывали PBS (Sigma-Aldrich, UK) три раза (10 минут / промывка). Ткани были проницаемы с помощью 0,1% Triton X-100 в PBS (Sigma-Aldrich, UK) в течение 30 минут, трижды промыли PBS (10 минут / промывка) и заблокировали 1% бычьим сывороточным альбумином (BSA; Sigma-Aldrich, UK). ) в PBS в течение ~ 30 мин.BSA удаляли, и срезы тканей инкубировали при 4 ° C в течение ночи с соответствующими первичными антителами, разведенными в 1% BSA в PBS. Все антитела, использованные в этом исследовании, были коммерчески доступны, проверены поставщиками на видовую реактивность и для иммунофлуоресценции и перечислены в дополнительной таблице 1. Были использованы разведения, рекомендованные поставщиком. После инкубации с первичными антителами в течение ночи срез ткани трижды промывали PBS (10 мин / промывка) и инкубировали в течение 2 ч при комнатной температуре с соответствующими вторичными антителами (см.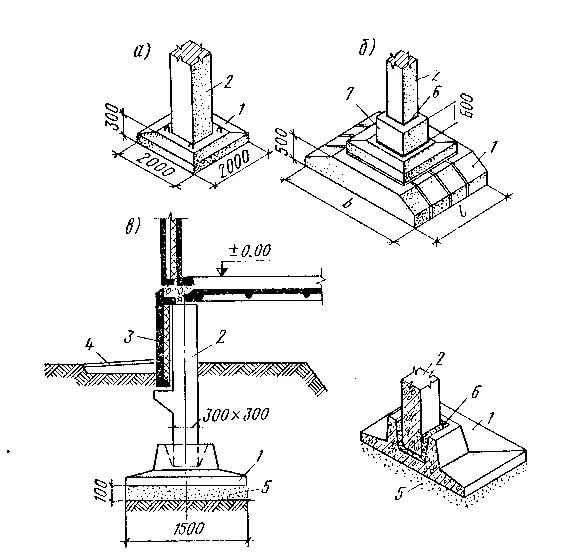 Таблицу со всей информацией об антителах в дополнительном материале), разведенными в 1% BSA в PBS.После инкубации со вторичными антителами срезы промывали трижды PBS (10 мин / промывка). Каплю монтажной среды Vectashield antitifade (H-1000, vector Laboratories, США) наносили на срезы тканей, а покровные стекла прикрепляли к предметным стеклам с лаком для ногтей. Иммунофлуоресцентное мечение белков детектировали с помощью лазерной сканирующей конфокальной микроскопии (LSM5, Carl Zeiss, Германия) с использованием программного обеспечения LSM5 Zeiss Pascal (Carl Zeiss, Германия) для получения изображений. Были использованы длины волн лазерного излучения и фильтры, подходящие для вторичных флуорофоров антител.Для каждого антитела использовали три среза ткани из каждого препарата правого предсердия, и из каждого среза ткани получали четыре изображения. Репрезентативные изображения мечения антител показаны на фиг. 1b, 4 и дополнительный рис. 17.
Таблицу со всей информацией об антителах в дополнительном материале), разведенными в 1% BSA в PBS.После инкубации со вторичными антителами срезы промывали трижды PBS (10 мин / промывка). Каплю монтажной среды Vectashield antitifade (H-1000, vector Laboratories, США) наносили на срезы тканей, а покровные стекла прикрепляли к предметным стеклам с лаком для ногтей. Иммунофлуоресцентное мечение белков детектировали с помощью лазерной сканирующей конфокальной микроскопии (LSM5, Carl Zeiss, Германия) с использованием программного обеспечения LSM5 Zeiss Pascal (Carl Zeiss, Германия) для получения изображений. Были использованы длины волн лазерного излучения и фильтры, подходящие для вторичных флуорофоров антител.Для каждого антитела использовали три среза ткани из каждого препарата правого предсердия, и из каждого среза ткани получали четыре изображения. Репрезентативные изображения мечения антител показаны на фиг. 1b, 4 и дополнительный рис. 17.
Регистрация внутриклеточного потенциала действия
Синусовый узел и потенциалы действия правого предсердия регистрировались с помощью острых микроэлектродов в препаратах спонтанно бьющихся тканей (рис. 1b). Раствор Тирода (содержание в мМ: NaCl 120,3, KCl 4,0, CaCl 2 1.2, MgSO 4 1,3, NaH 2 PO 4 1,2, NaHCO 3 25,2 и глюкоза 11) барботировали 95% O 2 /5% CO 2 Смесь для поддержания pH 7,4 циркулировала в 20 мл / мин, а ткани поддерживали при 37 ° C. Центр синусового узла был нанесен на карту с помощью набора из двух биполярных внеклеточных электродов. С помощью острых микроэлектродов, заполненных 3 M KCl, с сопротивлением кончика 20–40 МОм были получены множественные записи внутриклеточных потенциалов действия в центре синусового узла и гребешковой мышце (для ткани правого предсердия).Эти места записи соответствовали участкам биопсии ткани для MS. Разность напряжений на плазматической мембране регистрировалась на частоте 20 кГц, пропускалась через фильтр нижних частот Бесселя 10 кГц и усиливалась в 10 раз в усилителе Axon Instruments GeneClamp 500 (Molecular Devices, США). Данные были оцифрованы в Axon Instruments Digidata 1440A (Molecular Devices, США) и сохранены на компьютере для построения графиков потенциала действия (рис.
1b). Раствор Тирода (содержание в мМ: NaCl 120,3, KCl 4,0, CaCl 2 1.2, MgSO 4 1,3, NaH 2 PO 4 1,2, NaHCO 3 25,2 и глюкоза 11) барботировали 95% O 2 /5% CO 2 Смесь для поддержания pH 7,4 циркулировала в 20 мл / мин, а ткани поддерживали при 37 ° C. Центр синусового узла был нанесен на карту с помощью набора из двух биполярных внеклеточных электродов. С помощью острых микроэлектродов, заполненных 3 M KCl, с сопротивлением кончика 20–40 МОм были получены множественные записи внутриклеточных потенциалов действия в центре синусового узла и гребешковой мышце (для ткани правого предсердия).Эти места записи соответствовали участкам биопсии ткани для MS. Разность напряжений на плазматической мембране регистрировалась на частоте 20 кГц, пропускалась через фильтр нижних частот Бесселя 10 кГц и усиливалась в 10 раз в усилителе Axon Instruments GeneClamp 500 (Molecular Devices, США). Данные были оцифрованы в Axon Instruments Digidata 1440A (Molecular Devices, США) и сохранены на компьютере для построения графиков потенциала действия (рис. 3 вставки).
3 вставки).
Преобразование предсердия в SN-подобный потенциал действия модель
Может ли кардиостимуляция в синусовом узле быть объяснена различиями в экспрессии ионных каналов в синусовом узле? Чтобы ответить на этот вопрос, на основе данных масс-спектрометрии была модифицирована биофизически подробная модель потенциала действия предсердий мыши 14 .Во-первых, емкость ячейки была уменьшена с 50 до 25 пФ (типично для клеток предсердия и синусового узла соответственно). Количественные данные интенсивности без этикеток использовали для масштабирования проводимости ионного тока для I Na , I Ca, L и I K, r (дополнительная таблица 2). Другой подход должен быть использован для I f и I Ca, T , потому что они не присутствуют в модели потенциала действия предсердий мыши.Уравнения для I f и I Ca, T были взяты из независимой модели потенциала действия синусового узла мыши 15 и введены в модель потенциала действия предсердия мыши; для потенциала действия предсердий мыши ионные проводимости для двух токов были уменьшены в соответствии с данными масс-спектрометрии; при моделировании потенциала действия синусового узла ионные проводимости для двух токов не уменьшались. Кроме того, напряжение полуактивации I, , Ca, T было сдвинуто на -7 мВ по сравнению с образованием для I, , Ca, T из модели потенциала действия синусового узла мыши, из которой образовалось образование. принято 15 . I K, s не входит в модель предсердного потенциала действия мыши и, следовательно, не учитывалась. Ионные каналы (K v 1.5 и K v 4.x), отвечающие за I K, ur и I до , не были обнаружены. Тем не менее, ионные проводимости для I K, ur и I от до пришлось снизить до 0% и 10%, соответственно, потому что присутствие этих токов в синусовом узле привело к действию треугольной формы. потенциал; это следует рассматривать как ограничение моделирования, потому что это изменение при отсутствии обоснования со стороны масс-спектрометрии.Ионный канал, который обычно считается ответственным за I K, 1 (Kir2.
Кроме того, напряжение полуактивации I, , Ca, T было сдвинуто на -7 мВ по сравнению с образованием для I, , Ca, T из модели потенциала действия синусового узла мыши, из которой образовалось образование. принято 15 . I K, s не входит в модель предсердного потенциала действия мыши и, следовательно, не учитывалась. Ионные каналы (K v 1.5 и K v 4.x), отвечающие за I K, ur и I до , не были обнаружены. Тем не менее, ионные проводимости для I K, ur и I от до пришлось снизить до 0% и 10%, соответственно, потому что присутствие этих токов в синусовом узле привело к действию треугольной формы. потенциал; это следует рассматривать как ограничение моделирования, потому что это изменение при отсутствии обоснования со стороны масс-спектрометрии.Ионный канал, который обычно считается ответственным за I K, 1 (Kir2. x), не был обнаружен масс-спектрометрией. Возможно, что вместо этого за это ответственны каналы Kir3.x или TASK1, и оба ионных канала были менее многочисленными в синусовом узле (дополнительная таблица 2). Большинство расчетов проводилось в предположении, что I K, 1 не изменились в синусовом узле. Однако в некоторых моделях ионная проводимость для I K, 1 была уменьшена в соответствии с наблюдаемыми изменениями в Kir3.Каналы 1 и ТАСК1; это увеличивало частоту кардиостимуляции, но в остальном имело незначительный эффект (данные не показаны). Поглощение и высвобождение Ca 2+ в саркоплазматическом ретикулуме масштабировали в соответствии с данными масс-спектрометрии (дополнительная таблица 2). При моделировании модель потенциала действия предсердий стимулировалась с продолжительностью цикла 200 мс. Все клеточные модели были запущены в течение 20 с для достижения устойчивого состояния, и последние два потенциала действия были записаны для анализа.
x), не был обнаружен масс-спектрометрией. Возможно, что вместо этого за это ответственны каналы Kir3.x или TASK1, и оба ионных канала были менее многочисленными в синусовом узле (дополнительная таблица 2). Большинство расчетов проводилось в предположении, что I K, 1 не изменились в синусовом узле. Однако в некоторых моделях ионная проводимость для I K, 1 была уменьшена в соответствии с наблюдаемыми изменениями в Kir3.Каналы 1 и ТАСК1; это увеличивало частоту кардиостимуляции, но в остальном имело незначительный эффект (данные не показаны). Поглощение и высвобождение Ca 2+ в саркоплазматическом ретикулуме масштабировали в соответствии с данными масс-спектрометрии (дополнительная таблица 2). При моделировании модель потенциала действия предсердий стимулировалась с продолжительностью цикла 200 мс. Все клеточные модели были запущены в течение 20 с для достижения устойчивого состояния, и последние два потенциала действия были записаны для анализа. Характеристики смоделированных потенциалов действия показаны в дополнительной таблице 3.
Характеристики смоделированных потенциалов действия показаны в дополнительной таблице 3.
Мы сделали и обратное. Мы использовали модель Харче и соавт. 15 потенциала действия клеток синусового узла мыши в качестве отправной точки. Модель была изменена, чтобы включить дискретные ионные токи, переносимые различными каналами HCN. Затем модель была изменена таким же образом, как указано выше, но в обратном порядке. Чтобы успешно преобразовать потенциал действия синусового узла в потенциал действия, подобный предсердному (несамопроизвольный, треугольная форма потенциала действия), нам пришлось дополнительно изменить фоновый внутренний выпрямительный ток K + ( I К, 1 ).Поскольку масс-спектрометрия не предоставила данных о каналах Kir2 (ионные каналы, которые, как считается, отвечают за I K, 1 ), были включены I K, 1 из модели потенциала действия предсердных клеток мыши. Это привело к успешному преобразованию потенциала действия синусового узла в потенциал действия, подобный предсердному. Учитывая отсутствие протеомных доказательств для Kir2, можно было бы предположить, возможно ли, что не каналы Kir2 ответственны за I K, 1 , а вместо этого могут быть ответственны каналы TASK1 и / или Kir3, которые действительно изменяются в ожидаемом образом, если они несут ответственность за I K, 1 .
Учитывая отсутствие протеомных доказательств для Kir2, можно было бы предположить, возможно ли, что не каналы Kir2 ответственны за I K, 1 , а вместо этого могут быть ответственны каналы TASK1 и / или Kir3, которые действительно изменяются в ожидаемом образом, если они несут ответственность за I K, 1 .
Расчет количества ионных каналов в синусовом узле
Данные масс-спектрометрии были использованы для оценки относительного количества различных ионных каналов в миоците синусового узла. Чтобы преобразовать относительное количество ионных каналов в абсолютное количество ионных каналов в миоците синусового узла, требуется независимая оценка абсолютного количества ионных каналов в миоците синусового узла. Модель цепи Маркова может воспроизводить поведение одного ионного канала. Суммируя ионный ток для нескольких ионных каналов (все одного типа), можно оценить количество ионных каналов, необходимых для воспроизведения тока всей клетки. Модели цепи Маркова для пяти одиночных ионных каналов (Hcn1, Hcn4, Ca v 1.2, Ca v 3.2 и ERG) были разработаны на основе Wang et al. 83 модель потенциала действия синусового узла мыши. Эти модели цепи Маркова рассматривали открытое и закрытое состояние для Hcn1 и Hcn4 и два дополнительных неактивных состояния для Ca v 1.2, Ca v 3.2 и ERG (дополнительный рис. 9). Стохастическое поведение ионных каналов моделировалось с помощью метода Монте-Карло, имитирующего переход между состояниями.Используемые унитарные проводимости каналов перечислены в дополнительной таблице 4. Чтобы проверить стохастическое открытие и закрытие ионных каналов, мы смоделировали последовательные развертки моделей унитарных ионных каналов во время заранее заданного протокола фиксации напряжения (рис. 5 и дополнительные рис. 8). –11). Наконец, было вычислено количество каждого ионного канала, необходимое для учета всего тока ячейки. Рассчитанное количество ионных каналов HCN4 на миоцит синусового узла использовали для преобразования относительного количества ионных каналов в миоците синусового узла (на основании масс-спектрометрии) в абсолютное количество ионных каналов в миоците синусового узла.
Модели цепи Маркова для пяти одиночных ионных каналов (Hcn1, Hcn4, Ca v 1.2, Ca v 3.2 и ERG) были разработаны на основе Wang et al. 83 модель потенциала действия синусового узла мыши. Эти модели цепи Маркова рассматривали открытое и закрытое состояние для Hcn1 и Hcn4 и два дополнительных неактивных состояния для Ca v 1.2, Ca v 3.2 и ERG (дополнительный рис. 9). Стохастическое поведение ионных каналов моделировалось с помощью метода Монте-Карло, имитирующего переход между состояниями.Используемые унитарные проводимости каналов перечислены в дополнительной таблице 4. Чтобы проверить стохастическое открытие и закрытие ионных каналов, мы смоделировали последовательные развертки моделей унитарных ионных каналов во время заранее заданного протокола фиксации напряжения (рис. 5 и дополнительные рис. 8). –11). Наконец, было вычислено количество каждого ионного канала, необходимое для учета всего тока ячейки. Рассчитанное количество ионных каналов HCN4 на миоцит синусового узла использовали для преобразования относительного количества ионных каналов в миоците синусового узла (на основании масс-спектрометрии) в абсолютное количество ионных каналов в миоците синусового узла. Модели потенциалов действия решались численно с помощью программы для решения дифференциальных уравнений CVODE 84 . При моделировании использовался временной шаг 0,1 мс. Стохастическое поведение одноканальных моделей имитировалось методом Монте-Карло с меньшим временным шагом 0,02 мс. Мы рассчитали количество ионных каналов в миоцитах SN на основе их абсолютного количественного определения на основе интенсивности (iBAQ) 12 . Для каждого канального белка мы провели поиск в литературе, чтобы найти количество субъединиц в функциональном канале (например,g., четыре для HCN4 и один для Ca v 1.2) и разделили интенсивность iBAQ на количество субъединиц, составляющих канал (дополнительная таблица 3). Затем мы использовали нашу оценку 6255 каналов HCN4 на миоцит SN, полученную при моделировании цепи Маркова, для преобразования интенсивности iBAQ в число копий на клетку. На основе этих данных мы рассчитали среднее значение и стандартную ошибку среднего, которые представлены в виде гистограмм (рис.
Модели потенциалов действия решались численно с помощью программы для решения дифференциальных уравнений CVODE 84 . При моделировании использовался временной шаг 0,1 мс. Стохастическое поведение одноканальных моделей имитировалось методом Монте-Карло с меньшим временным шагом 0,02 мс. Мы рассчитали количество ионных каналов в миоцитах SN на основе их абсолютного количественного определения на основе интенсивности (iBAQ) 12 . Для каждого канального белка мы провели поиск в литературе, чтобы найти количество субъединиц в функциональном канале (например,g., четыре для HCN4 и один для Ca v 1.2) и разделили интенсивность iBAQ на количество субъединиц, составляющих канал (дополнительная таблица 3). Затем мы использовали нашу оценку 6255 каналов HCN4 на миоцит SN, полученную при моделировании цепи Маркова, для преобразования интенсивности iBAQ в число копий на клетку. На основе этих данных мы рассчитали среднее значение и стандартную ошибку среднего, которые представлены в виде гистограмм (рис. 5 и дополнительный рис. 7).
5 и дополнительный рис. 7).
Электронная микроскопия
Образцы ткани синусового узла и правого предсердия (~ 1 × 1 мм) собирали, как описано выше для масс-спектрометрии.Протокол от Starborg et al. 85 был использован для подготовки и изображения образцов ткани для электронной микроскопии. В частности, свеже собранные образцы фиксировали 2,5% глутаровым альдегидом в 100 мМ какодилатном буфере (pH 7,2) в течение не менее 1 часа и хранили при 4 ° C до дальнейшего использования. Ткани трижды промывали дистиллированной водой (5 мин / промывка) и помещали в свежеприготовленную смесь 1% тетроксида осмия и 1,5% ферроцианида калия в 0,1 М какодилатном буфере на 1-2 часа. Затем ткани трижды промывали дистиллированной водой (5 мин / промывка) и переносили в свежеприготовленную 1% дубильную кислоту в 0.1 M какодилатный буфер в течение 1-2 часов с последующими пятью промывками дистиллированной водой (5 минут / промывка). Очень важно тщательно смыть дубильную кислоту. Образцы инкубировали в 1% -ном четырехокиси осмия в бидистиллированной воде в течение 40 мин при комнатной температуре, а затем промывали дистиллированной водой (три промывки, 5 мин / промывка) перед инкубацией в водном 1% -ном уранилацетате в течение не менее 1 ч и хранили в течение ночи при температуре окружающей среды. 4 ° С. На следующий день образцы трижды промывали дистиллированной водой (5 мин / промывка) и обезвоживали путем последующих замен следующих разведений этанола в дистиллированной воде: 30, 50, 70, 90 и два раза 100% по 15 минут каждый раз. шаг при комнатной температуре.Затем следовали две смены ацетона по 30 мин каждая. Образцы пропитывали серией градуированных смол TAAB 812 Hard (TAAB Laboratories Equipment Ltd, UK) в ацетоне при комнатной температуре: сначала в течение ночи в 25% TAAB 812, затем в течение 8 часов в 50% TAAB 812, а затем в течение ночи в 75% TAAB 812. и, наконец, 6 часов в 100% TAAB. Образцы переносили на свежий 100% TAAB 812 Hard в маркированных формах и оставляли для отверждения при 60 ° C в течение 24–48 часов. Небольшие кусочки залитых смолой тканей закрепляли на алюминиевых шпильках для образцов (Gatan, США) с помощью цианоакрилатного клея.Блоки были облицованы и прецизионно обрезаны алмазным ножом до квадрата (~ 0,5 × 0,5 мм), так что ткань обнажалась со всех четырех сторон, и подготовлены для исследования с помощью микротома Gatan 3view (Gatan, США) внутри FEI Quanta 250 FEG.
4 ° С. На следующий день образцы трижды промывали дистиллированной водой (5 мин / промывка) и обезвоживали путем последующих замен следующих разведений этанола в дистиллированной воде: 30, 50, 70, 90 и два раза 100% по 15 минут каждый раз. шаг при комнатной температуре.Затем следовали две смены ацетона по 30 мин каждая. Образцы пропитывали серией градуированных смол TAAB 812 Hard (TAAB Laboratories Equipment Ltd, UK) в ацетоне при комнатной температуре: сначала в течение ночи в 25% TAAB 812, затем в течение 8 часов в 50% TAAB 812, а затем в течение ночи в 75% TAAB 812. и, наконец, 6 часов в 100% TAAB. Образцы переносили на свежий 100% TAAB 812 Hard в маркированных формах и оставляли для отверждения при 60 ° C в течение 24–48 часов. Небольшие кусочки залитых смолой тканей закрепляли на алюминиевых шпильках для образцов (Gatan, США) с помощью цианоакрилатного клея.Блоки были облицованы и прецизионно обрезаны алмазным ножом до квадрата (~ 0,5 × 0,5 мм), так что ткань обнажалась со всех четырех сторон, и подготовлены для исследования с помощью микротома Gatan 3view (Gatan, США) внутри FEI Quanta 250 FEG.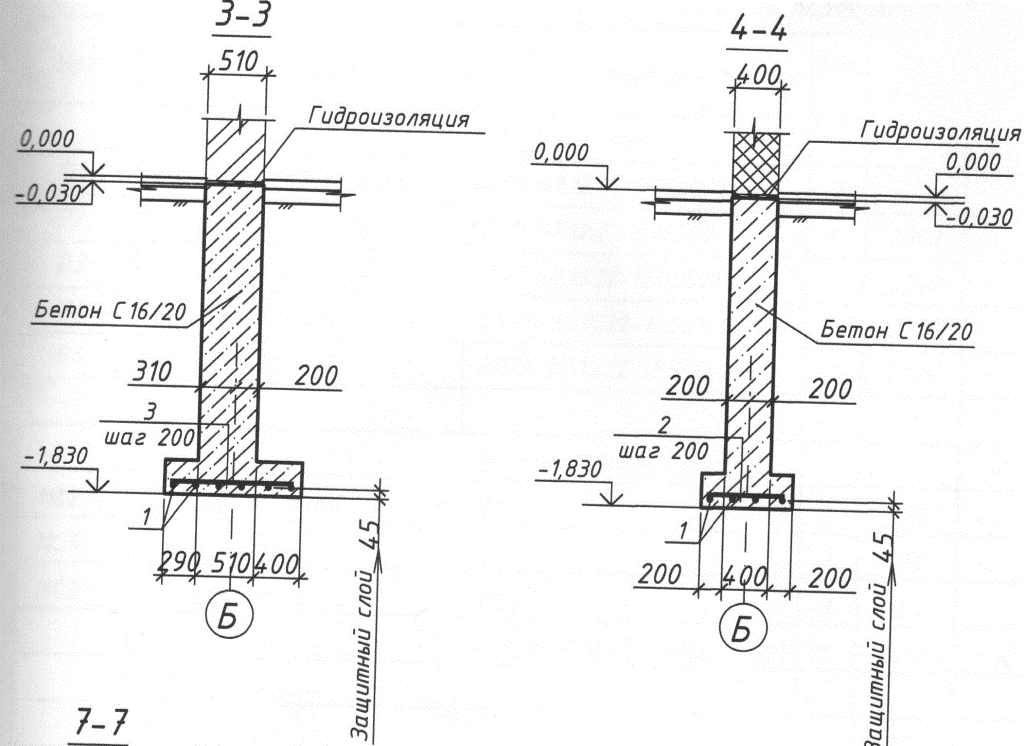 (Thermo Fisher Scientific, Великобритания) серийный блок-сканирующий электрон. Последовательные изображения поверхности блока были получены при ускоряющем напряжении 3,8 кВ, размере пятна 2,8 и давлении 1,6 торр. Время выдержки для каждой микрофотографии составляло 4 мкс. Размер пикселя 24,4 × 24.4 нм (XxY), а толщина Z-среза составляла 75 нм. Ширина поля по горизонтали и вертикали составляла ~ 100 и ~ 200 мкм соответственно. Для просвечивающей электронной микроскопии (ПЭМ) срезы вырезали на ультрамикротоме Reichert Ultracut и наблюдали с помощью микроскопа FEI Tecnai 12 BioTWIN (FEI Company, США) при ускоряющем напряжении 100 кВ. Изображения были получены с помощью камеры Gatan Orius SC1000 CCD и представлены на дополнительных рисунках. 14–16.
(Thermo Fisher Scientific, Великобритания) серийный блок-сканирующий электрон. Последовательные изображения поверхности блока были получены при ускоряющем напряжении 3,8 кВ, размере пятна 2,8 и давлении 1,6 торр. Время выдержки для каждой микрофотографии составляло 4 мкс. Размер пикселя 24,4 × 24.4 нм (XxY), а толщина Z-среза составляла 75 нм. Ширина поля по горизонтали и вертикали составляла ~ 100 и ~ 200 мкм соответственно. Для просвечивающей электронной микроскопии (ПЭМ) срезы вырезали на ультрамикротоме Reichert Ultracut и наблюдали с помощью микроскопа FEI Tecnai 12 BioTWIN (FEI Company, США) при ускоряющем напряжении 100 кВ. Изображения были получены с помощью камеры Gatan Orius SC1000 CCD и представлены на дополнительных рисунках. 14–16.
Протеомные измерения фибробластов
Сердца были эксплантированы от трех мышей-самцов, промыты ледяным фосфатно-солевым буфером (PBS) и желудочки рассечены на льду.Желудочки были тонко разрезаны скальпелем и переварены с использованием 2,5 мл коктейля для переваривания (Miltenyi Biotech) с последующим растиранием.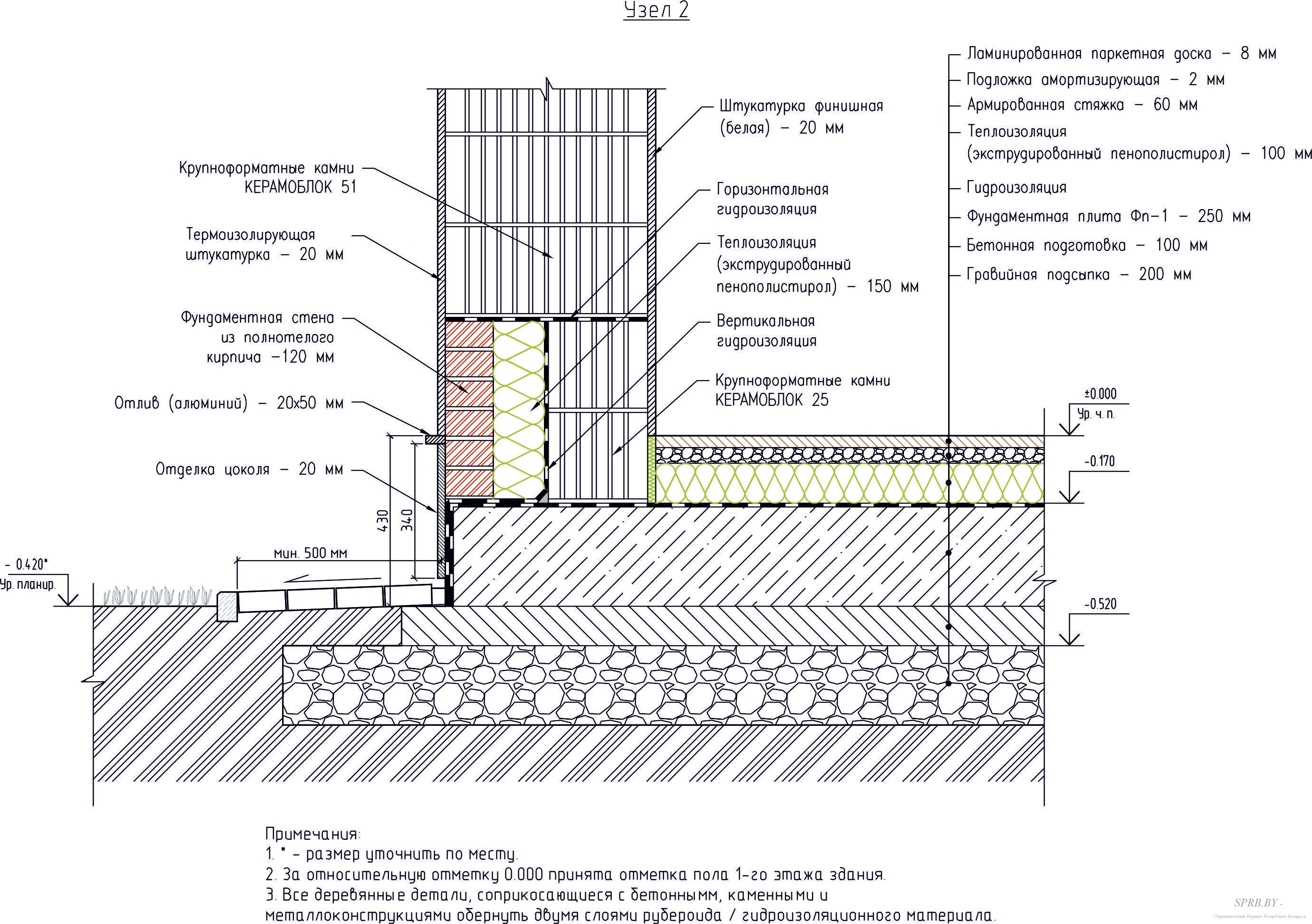 Выделение DMEM [с высоким содержанием глюкозы, глутамакса, без пирувата, 100 мкм аскорбиновой кислоты, 20% фетальной телячьей сыворотки (FBS, Gibco), 0–9% пенициллина / стрептомицина (P / S, Gibco)] добавляли к каждому образцу с последующей фильтрацией через сетчатый фильтр для клеток 70 мкм. Фильтраты центрифугировали при 500 × g в течение 5 мин, осадок клеток ресуспендировали в 1 мл PEB-буфера (фосфатно-солевой буферный раствор (PBS), pH 7.2, 0,5% бычьего сывороточного альбумина (BSA), 2 мМ EDTA). Добавляли 10 мл 1x раствора для лизиса эритроцитов (Miltenyi Biotech) и образцы центрифугировали при 600 × g в течение 5 мин. Каждый осадок клеток ресуспендировали в PBS, содержащем 15 мкл фермента A (Miltenyi Biotech), и центрифугировали при 600 × g в течение 5 мин. Осадки клеток ресуспендировали в [с высоким содержанием глюкозы, глутамакса, без пирувата, 100 мкм аскорбиновой кислоты, 9% FBS, 0–9% P / S] и высевали на чашки, покрытые поли-d-лизином. Через 2 часа при 37 ° C в увлажненном инкубаторе с 5% CO2 клетки дважды промывали, отделяли и подсчитывали, используя анализ жизнеспособности и подсчета клеток на Nucleocounter 3000 (Chemometec).
Выделение DMEM [с высоким содержанием глюкозы, глутамакса, без пирувата, 100 мкм аскорбиновой кислоты, 20% фетальной телячьей сыворотки (FBS, Gibco), 0–9% пенициллина / стрептомицина (P / S, Gibco)] добавляли к каждому образцу с последующей фильтрацией через сетчатый фильтр для клеток 70 мкм. Фильтраты центрифугировали при 500 × g в течение 5 мин, осадок клеток ресуспендировали в 1 мл PEB-буфера (фосфатно-солевой буферный раствор (PBS), pH 7.2, 0,5% бычьего сывороточного альбумина (BSA), 2 мМ EDTA). Добавляли 10 мл 1x раствора для лизиса эритроцитов (Miltenyi Biotech) и образцы центрифугировали при 600 × g в течение 5 мин. Каждый осадок клеток ресуспендировали в PBS, содержащем 15 мкл фермента A (Miltenyi Biotech), и центрифугировали при 600 × g в течение 5 мин. Осадки клеток ресуспендировали в [с высоким содержанием глюкозы, глутамакса, без пирувата, 100 мкм аскорбиновой кислоты, 9% FBS, 0–9% P / S] и высевали на чашки, покрытые поли-d-лизином. Через 2 часа при 37 ° C в увлажненном инкубаторе с 5% CO2 клетки дважды промывали, отделяли и подсчитывали, используя анализ жизнеспособности и подсчета клеток на Nucleocounter 3000 (Chemometec).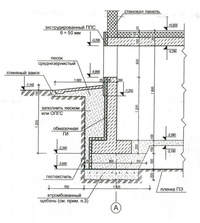 Клетки центрифугировали при 10000 × г в течение 7 минут и после удаления супернатанта замораживали при -80 ° C. Для получения пептида три образца фибробластов обрабатывали отдельно. Сначала клетки лизировали, добавляя 20 мкл 1% раствора TritonX-100 (1x полный ингибитор протеиназы (Roche), 50 мМ β-глицерофосфат, 10 мМ ортованадат натрия, 5 мМ хлорид магния в PBS). Добавляли один микрограмм ДНКазы, и образцы инкубировали на льду в течение 1 ч при встряхивании. 70 мкг гранул (1: 1 запас магнитных частиц, модифицированных карбоксилатом Sera-Mag (гидрофильные, 24152105050250) и магнитных частиц, модифицированных карбоксилатом Sera-Mag (гидрофобные 44152105050250 от GE Healthcare), добавляли к каждому образцу, и этанол добавляли к конечной стадии 70%.Через 10 минут при комнатной температуре (RT) образцы помещали на магнит на 1 минуту, шарики дважды промывали 70% этанолом и затем ресуспендировали в 20 мкл 50 мМ HEPES pH 8,5. TCEP добавляли до конечной концентрации 5 мМ и CAA до конечной концентрации 5,5 мМ.
Клетки центрифугировали при 10000 × г в течение 7 минут и после удаления супернатанта замораживали при -80 ° C. Для получения пептида три образца фибробластов обрабатывали отдельно. Сначала клетки лизировали, добавляя 20 мкл 1% раствора TritonX-100 (1x полный ингибитор протеиназы (Roche), 50 мМ β-глицерофосфат, 10 мМ ортованадат натрия, 5 мМ хлорид магния в PBS). Добавляли один микрограмм ДНКазы, и образцы инкубировали на льду в течение 1 ч при встряхивании. 70 мкг гранул (1: 1 запас магнитных частиц, модифицированных карбоксилатом Sera-Mag (гидрофильные, 24152105050250) и магнитных частиц, модифицированных карбоксилатом Sera-Mag (гидрофобные 44152105050250 от GE Healthcare), добавляли к каждому образцу, и этанол добавляли к конечной стадии 70%.Через 10 минут при комнатной температуре (RT) образцы помещали на магнит на 1 минуту, шарики дважды промывали 70% этанолом и затем ресуспендировали в 20 мкл 50 мМ HEPES pH 8,5. TCEP добавляли до конечной концентрации 5 мМ и CAA до конечной концентрации 5,5 мМ. Через 30 минут при комнатной температуре добавляли 0,25 мкг лиз-С (Trichem ApS, Дания) и образцы инкубировали при 37 ° C в течение 1 часа. Кроме того, добавляли 0,5 мкг трипсина (Life Technologies, США) и образцы расщепляли в течение ночи при 37 ° C. Пептиды отделяли от шариков с помощью магнитного поля, переносили в новую пробирку и подкисляли до конечной концентрации 1% трифторуксусной кислоты (TFA).Пептиды очищали с использованием картриджей C 18 SepPak (Waters). Пептиды элюировали 40% ацетонитрилом, содержащим 0,05% TFA. Перед измерением методом масс-спектрометрии обессоленную смесь пептидов разделяли на 12 конкатенированных фракций в системе Dionex UltiMate 3000 UPLC (Thermo Scientific, США), как описано выше 6,7 . Образцы фракционированных пептидов вводили в колонку 15 см (внутренний диаметр 75 мкМ), заполненную шариками C18 1,9 мкМ (Dr. Maisch GmbH), используя Easy-LC 1200 (Thermo Fisher Scientific), и разделяли в 1-часовом линейном градиенте с увеличивающимся буфером.
Через 30 минут при комнатной температуре добавляли 0,25 мкг лиз-С (Trichem ApS, Дания) и образцы инкубировали при 37 ° C в течение 1 часа. Кроме того, добавляли 0,5 мкг трипсина (Life Technologies, США) и образцы расщепляли в течение ночи при 37 ° C. Пептиды отделяли от шариков с помощью магнитного поля, переносили в новую пробирку и подкисляли до конечной концентрации 1% трифторуксусной кислоты (TFA).Пептиды очищали с использованием картриджей C 18 SepPak (Waters). Пептиды элюировали 40% ацетонитрилом, содержащим 0,05% TFA. Перед измерением методом масс-спектрометрии обессоленную смесь пептидов разделяли на 12 конкатенированных фракций в системе Dionex UltiMate 3000 UPLC (Thermo Scientific, США), как описано выше 6,7 . Образцы фракционированных пептидов вводили в колонку 15 см (внутренний диаметр 75 мкМ), заполненную шариками C18 1,9 мкМ (Dr. Maisch GmbH), используя Easy-LC 1200 (Thermo Fisher Scientific), и разделяли в 1-часовом линейном градиенте с увеличивающимся буфером.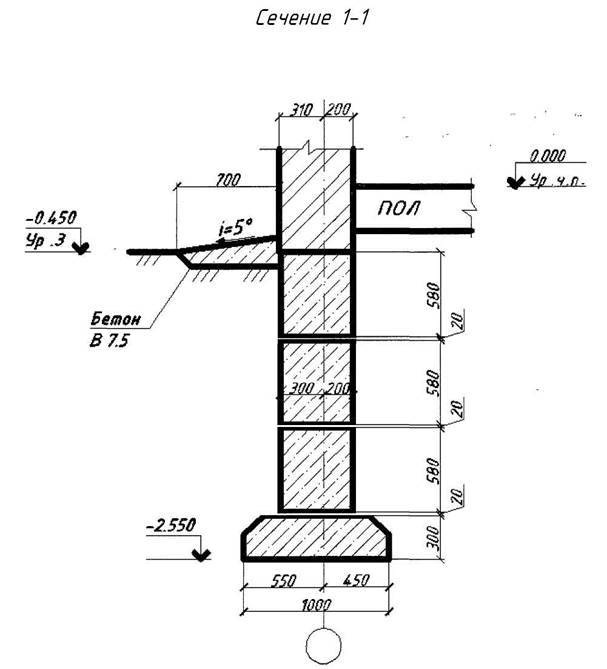 B (80% ацетонитрил, 0.1% муравьиной кислоты) при расходе 250 нл / мин. Все образцы анализировали на масс-спектрометре Q-Exactive HF-X (Thermo Fisher Scientific), работающем в положительном, зависимом от данных режиме, с использованием метода Top12, позволяющего проводить глубокие измерения протеома (разрешение MS1 = 60,000, разрешение MS2 = 15,000). Необработанные данные МС обрабатывали с использованием программного обеспечения MaxQuant v1.6.1.0 77 с использованием настроек по умолчанию и базы данных, содержащей все проверенные записи белков SwissProt (загружены 26 июля 2018 г.).
B (80% ацетонитрил, 0.1% муравьиной кислоты) при расходе 250 нл / мин. Все образцы анализировали на масс-спектрометре Q-Exactive HF-X (Thermo Fisher Scientific), работающем в положительном, зависимом от данных режиме, с использованием метода Top12, позволяющего проводить глубокие измерения протеома (разрешение MS1 = 60,000, разрешение MS2 = 15,000). Необработанные данные МС обрабатывали с использованием программного обеспечения MaxQuant v1.6.1.0 77 с использованием настроек по умолчанию и базы данных, содержащей все проверенные записи белков SwissProt (загружены 26 июля 2018 г.).
Одноядерное секвенирование РНК
Синусовые узлы выделяли у 16 мышей, как описано выше.Синусные узлы четырех животных были объединены, быстро заморожены в жидком азоте и хранились при -80 ° C до обработки. Для выделения ядер образцы хранили на сухом льду перед добавлением 1 мл буфера для лизиса Nuclei EZ (Sigma-Aldrich). Ткань растирали пипеткой 10х с наконечником на 1000 мкл, а затем 10х с наконечником на 200 мкл.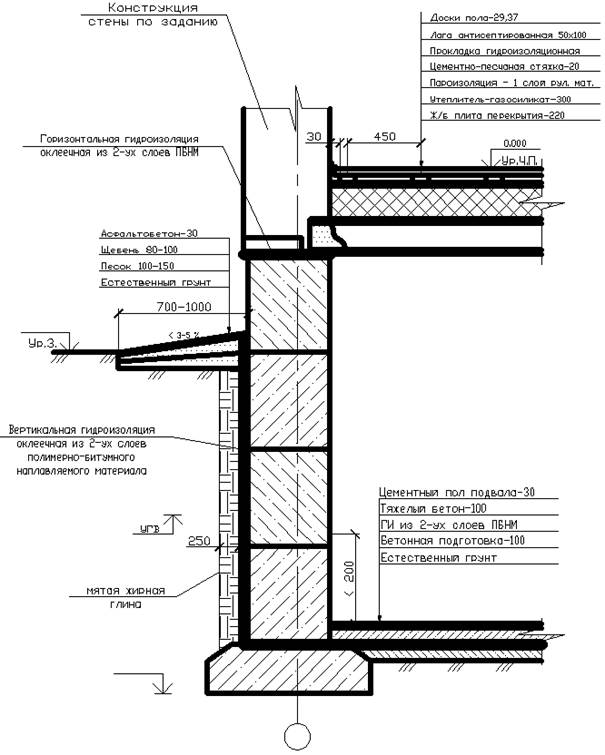 Образцы инкубировали на льду в течение 5 мин, переворачивая пробирки каждую минуту. После еще одного цикла растирания (наконечник 10 × 1000 мкл) было проведено центрифугирование при 800 × g в течение 7 мин.Супернатант осторожно удаляли, а ядра и оставшиеся кусочки ткани ресуспендировали в 500 мкл буфера для ресуспендирования (0,1% BSA, содержащий 0,2 ед. / Мкл ингибитора протекторной РНКазы). Образцы фильтровали через нейлоновую сетку 70 мкм, которая была пропитана РНКазой в течение ночи и промывалась в MilliQ. После этого фильтр промывали 500 мкл буфера для ресуспендирования и образцы центрифугировали при 800 × g в течение 7 мин. Девятьсот девяносто микролитров супернатанта удаляли и добавляли 10 мкл буфера для ресуспендирования.Выделенные ядра были визуально проверены и загружены в два канала системы Chromium (10x геномика) с использованием протокола производителя (Chromium Single-Cell 3 ‘Reagent Kits (v3 Chemistry)) для получения одноядерных библиотек, которые были секвенированы на NovaSeq ™ 6000 система секвенирования (Illumina) с использованием набора реагентов S1 (100 циклов).
Образцы инкубировали на льду в течение 5 мин, переворачивая пробирки каждую минуту. После еще одного цикла растирания (наконечник 10 × 1000 мкл) было проведено центрифугирование при 800 × g в течение 7 мин.Супернатант осторожно удаляли, а ядра и оставшиеся кусочки ткани ресуспендировали в 500 мкл буфера для ресуспендирования (0,1% BSA, содержащий 0,2 ед. / Мкл ингибитора протекторной РНКазы). Образцы фильтровали через нейлоновую сетку 70 мкм, которая была пропитана РНКазой в течение ночи и промывалась в MilliQ. После этого фильтр промывали 500 мкл буфера для ресуспендирования и образцы центрифугировали при 800 × g в течение 7 мин. Девятьсот девяносто микролитров супернатанта удаляли и добавляли 10 мкл буфера для ресуспендирования.Выделенные ядра были визуально проверены и загружены в два канала системы Chromium (10x геномика) с использованием протокола производителя (Chromium Single-Cell 3 ‘Reagent Kits (v3 Chemistry)) для получения одноядерных библиотек, которые были секвенированы на NovaSeq ™ 6000 система секвенирования (Illumina) с использованием набора реагентов S1 (100 циклов).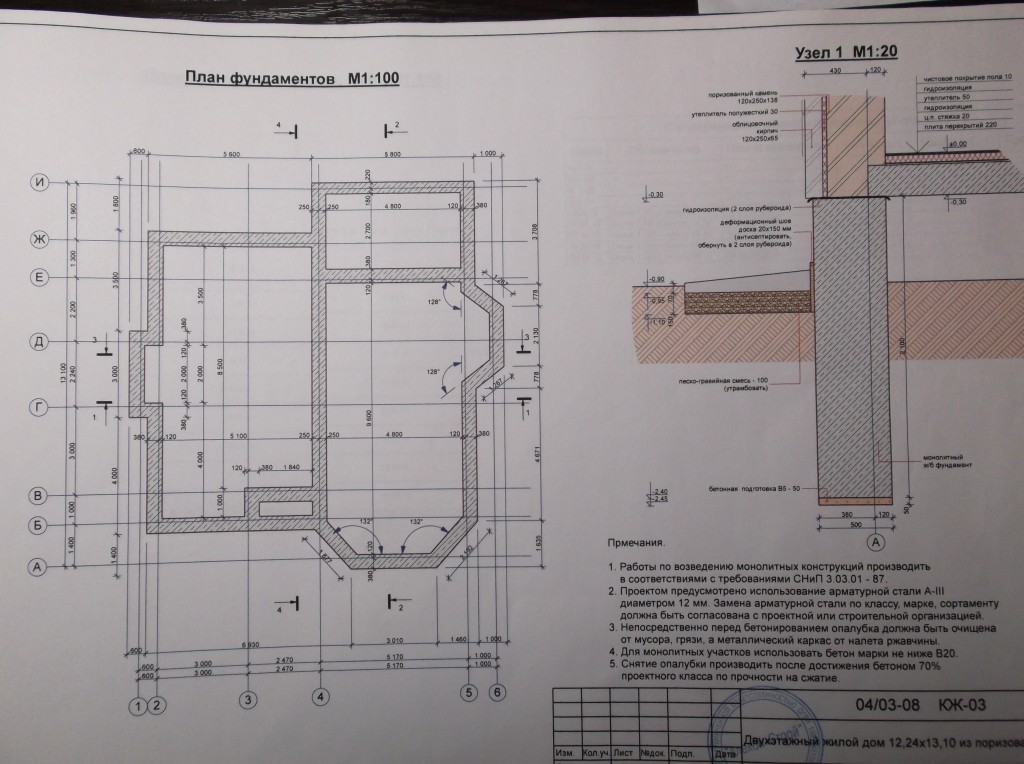 Для выравнивания считывания при секвенировании одноядерной РНК использовался конвейер Cellranger 3.0 (10x Genomics, США) для сопоставления считываний, количественной оценки уникальных молекулярных идентификаторов (UMI) и создания матриц экспрессии штрих-кода признаков.Считывания были сопоставлены с адаптированным эталоном из 10x mm10-premrna для включения интронов в соответствии с процедурой, изложенной на веб-сайте 10x Genomics [https://support.10xgenomics.com/single-cell-gene-expression/software/pipelines/ последние / расширенные / ссылки]. Необработанные матрицы экспрессии, полученные с помощью инструмента подсчета Cellranger, впоследствии были проанализированы с использованием пакета Seurat v.3.0.0.9000 R 86 . В соответствии с показаниями 10x Genomics, для каждого образца, исходя из примерно 9000 загруженных клеток, для последующего анализа были первоначально выбраны 5157 штрих-кодов с наибольшим числом отсчетов [https: // kb.10xgenomics.com/hc/en-us/articles/360001378811-What-is-the-maximum-number-of-cells-that-can-be-profiled-].
Для выравнивания считывания при секвенировании одноядерной РНК использовался конвейер Cellranger 3.0 (10x Genomics, США) для сопоставления считываний, количественной оценки уникальных молекулярных идентификаторов (UMI) и создания матриц экспрессии штрих-кода признаков.Считывания были сопоставлены с адаптированным эталоном из 10x mm10-premrna для включения интронов в соответствии с процедурой, изложенной на веб-сайте 10x Genomics [https://support.10xgenomics.com/single-cell-gene-expression/software/pipelines/ последние / расширенные / ссылки]. Необработанные матрицы экспрессии, полученные с помощью инструмента подсчета Cellranger, впоследствии были проанализированы с использованием пакета Seurat v.3.0.0.9000 R 86 . В соответствии с показаниями 10x Genomics, для каждого образца, исходя из примерно 9000 загруженных клеток, для последующего анализа были первоначально выбраны 5157 штрих-кодов с наибольшим числом отсчетов [https: // kb.10xgenomics.com/hc/en-us/articles/360001378811-What-is-the-maximum-number-of-cells-that-can-be-profiled-].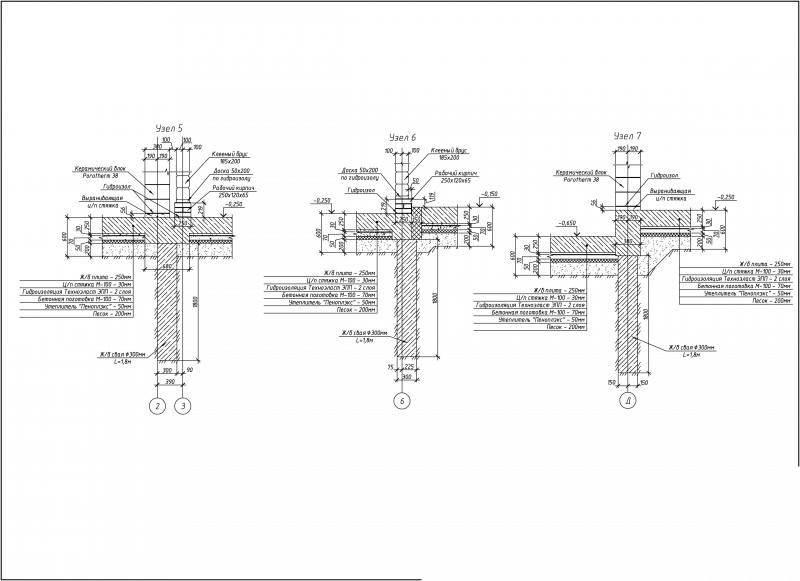 На следующем этапе фильтрации, основанном на визуальном осмотре графиков контроля качества, были сохранены штрих-коды с не менее 3000 UMI и пропорциями митохондриальной и рибосомной РНК ниже 0,08 и 0,1 соответственно. Пакет DoubletFinder R 87 использовался для удаления подозреваемых дуплетов. Из исходных 5127 штрих-кодов, выбранных из каждого канала, 4426 ядер из первого канала и 931 из второго канала пережили QC, причем максимальные значения экспрессии митохрондриальной и рибосомной экспрессии составляют большинство пропущенных штрих-кодов.Данные были нормализованы с помощью команды LogNormalize (), команда FindVariableFeatures () использовалась для поиска верхних 2000 генов переменных, а команда ScaleData () использовалась для масштабирования данных экспрессии до баллов Z и регрессии сопутствующих вариаций, связанных с с числом UMI или с содержанием рибосомных или митохондриальных генов. Команда RunPCA () использовалась для вычисления 75 основных компонентов (ПК).
На следующем этапе фильтрации, основанном на визуальном осмотре графиков контроля качества, были сохранены штрих-коды с не менее 3000 UMI и пропорциями митохондриальной и рибосомной РНК ниже 0,08 и 0,1 соответственно. Пакет DoubletFinder R 87 использовался для удаления подозреваемых дуплетов. Из исходных 5127 штрих-кодов, выбранных из каждого канала, 4426 ядер из первого канала и 931 из второго канала пережили QC, причем максимальные значения экспрессии митохрондриальной и рибосомной экспрессии составляют большинство пропущенных штрих-кодов.Данные были нормализованы с помощью команды LogNormalize (), команда FindVariableFeatures () использовалась для поиска верхних 2000 генов переменных, а команда ScaleData () использовалась для масштабирования данных экспрессии до баллов Z и регрессии сопутствующих вариаций, связанных с с числом UMI или с содержанием рибосомных или митохондриальных генов. Команда RunPCA () использовалась для вычисления 75 основных компонентов (ПК).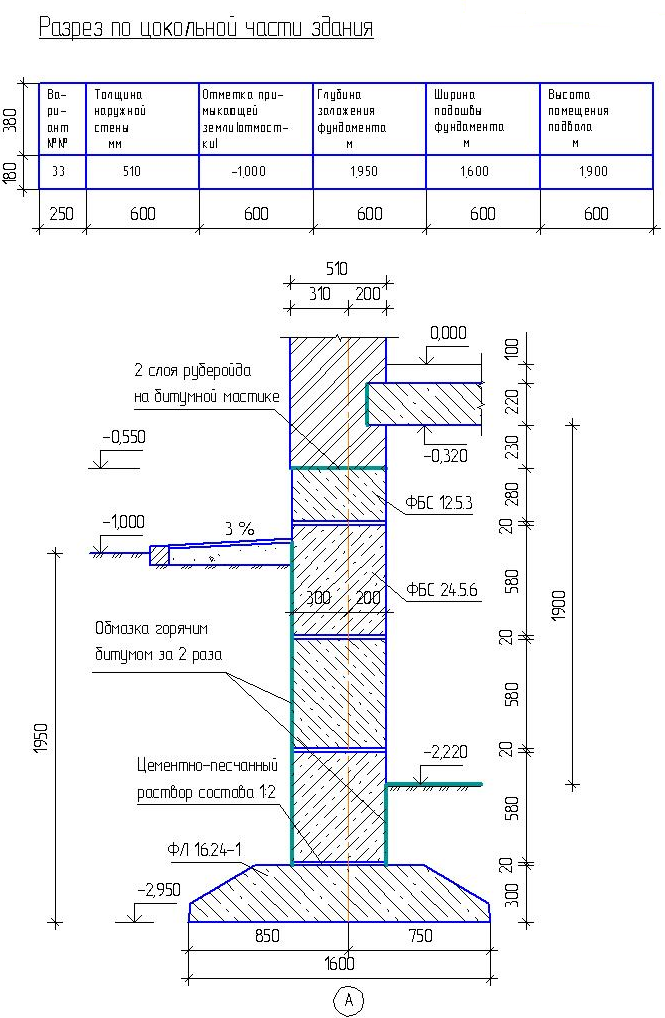 Команды FindNeighbors () и FindClusters () использовались с параметрами по умолчанию для идентификации кластеров ячеек, а команда FindMarkers () использовалась с методом MAST из пакета MAST R 88 (v1.8.2, https://github.com/RGLab/MAST/) для идентификации дифференциально экспрессируемых генов. Эти гены использовались для ручного добавления аннотаций типов клеток в кластеры. Команда RunTSNE () использовалась с точностью 30, чтобы произвести 2-D T-распределенное стохастическое вложение соседей (t-SNE). Кластеры и панели экспрессии генов визуализировали с использованием пакета ggplot2 R 89 .
Команды FindNeighbors () и FindClusters () использовались с параметрами по умолчанию для идентификации кластеров ячеек, а команда FindMarkers () использовалась с методом MAST из пакета MAST R 88 (v1.8.2, https://github.com/RGLab/MAST/) для идентификации дифференциально экспрессируемых генов. Эти гены использовались для ручного добавления аннотаций типов клеток в кластеры. Команда RunTSNE () использовалась с точностью 30, чтобы произвести 2-D T-распределенное стохастическое вложение соседей (t-SNE). Кластеры и панели экспрессии генов визуализировали с использованием пакета ggplot2 R 89 .
Сводка отчетов
Дополнительная информация о дизайне исследования доступна в Сводке отчетов по исследованиям природы, связанной с этой статьей.
Фундамент | SKALE
Фонд Сети децентрализованной экономики (N.O.D.E.), базирующийся в Лихтенштейне, был создан для выполнения миссии поддержки сети SKALE. Сеть SKALE предназначена для поддержки предприятий, людей и организаций, работающих в открытом Интернете.
Сеть SKALE в конечном итоге регулируется своим кодом, который включает токен SKL. Ончейн-голосование потребуется для изменения любых экономических принципов протокола.
Фонд будет поддерживать сеть путем избрания разнообразного набора сетевых представителей, состоящих из разработчиков Dapp, которые запускают SKALE Chains, валидаторов, которые запускают узлы SKALE, независимых, таких как инвесторы, которые помогли запустить экономику сети, и разработчиков / представителей, которые активно создают, поддерживать и развивать кодовую базу.Эти представители будут служить сообществу, содействуя голосованию в сети, грантам и решениям по бюджету / казначейству. Первоначально представители будут выбраны Фондом, однако будущие выборы будут проводиться с помощью варианта голосования держателей токенов в сети, чтобы выбрать представителей, которые облегчат голосование в сети.
Чтобы обеспечить долговечность Фонда, ему было выделено 10 процентов от общего пула токенов SKL, передаваемых на семилетний период, чтобы обеспечить его бюджетом и ресурсами для выполнения своей миссии.
Перспективный путь для SKALE Labs
SKALE Labs передала фонду активы интеллектуальной собственности и средства, собранные посредством первоначальных SAFT. Компания SKALE заключила договор на обслуживание с N.O.D.E. Anstalt предоставляет вклады кода в репозиторий с открытым исходным кодом, маркетинговую поддержку, поддержку построения сообщества, разработку решений и другие положительные вклады сообщества в сеть. SKALE по-прежнему будет регулярно вносить свой вклад в репозитории сети SKALE и поддерживать его, а также развивать и поддерживать сообщество.Эта роль не включает контроль или управление. SKALE Labs, как и любая другая организация, является сторонником сети извне.
Контракт SKALE Labs будет контролироваться и управляться представителями сети Фонда для обеспечения децентрализации, а также того, чтобы цели и видение сообщества всегда становились высшим приоритетом.
обновлений группы разработчиков Node.js. Поскольку в Node.js столько всего происходит… | автор: Node.
 js
jsВ связи с тем, что в сообществе Node.js и с Node.js Foundation, мы хотели воспользоваться моментом, чтобы напомнить сообществу, кто из Linux Foundation что делает для поддержки Node.js Foundation. Ниже приводится обновленная информация:
Майк Долан, исполняющий обязанности исполнительного директора (GitHub: @mkdolan, [email protected], Twitter: @mdolan
Майк является временным исполнительным директором фонда Node.js и вице-президентом по стратегическим программам поддержки проекты с открытым исходным кодом и юридические программы в Linux Foundation
Кейтлин Барнард, Node.js Foundation Marketing (GitHub: kbarnard10, [email protected] Twitter: @ kaitlyn_barnard)
Кейтлин уже курирует контент для JS Devs Zone JS Foundation, ежемесячного информационного бюллетеня и взаимодействия с участниками. Она присоединилась к команде Node.js Foundation, чтобы отвечать за ежемесячную рассылку новостей и обновления веб-сайтов. Она также будет тесно сотрудничать с Рэйчел Ромофф в области контент-маркетинга.
Она также будет тесно сотрудничать с Рэйчел Ромофф в области контент-маркетинга.
Аманда Эннис, социальные сети фонда Node.js (GitHub: OfficialLinuxAmanda, aennis @ linuxfoundation.org, Twitter: @ Linux_Amanda )
Аманда имеет более чем 6-летний опыт работы с социальными сетями, как платными, так и обычными. Аманда будет работать в социальных сетях и сотрудничать с маркетинговой командой и сообществом Node.js. над контентом.
Рэйчел Ромофф, по связям с общественностью фонда Node.js (Github: @Rromoff, [email protected], Twitter: @rachelromoff)
В качестве менеджера по связям с общественностью в Node.js Foundation Рэйчел продолжит создавать и выполнять коммуникационные программы, в том числе возможности выступлений, отношения со СМИ и курирование контента через Node.js Foundation Blog и сборник Node.js. Она является основной частью команды Node.js по социальным сетям и цифровому маркетингу.
Брайан Уорнер, Node. js Foundation Operations (GitHub: brianwarner, [email protected], Twitter: @realbrianwarner)
js Foundation Operations (GitHub: brianwarner, [email protected], Twitter: @realbrianwarner)
Брайан отвечает за текущие операции. Это включает в себя как повседневную работу, так и обеспечение того, чтобы логистические аспекты JSF и Node.js Foundation были готовы к слиянию.
Сара Конвей (Github: @SarahKConway, sconway @ linuxfoundation.org, Twitter: @SarahKConway) продолжает работать в качестве старшего руководителя маркетинговых коммуникаций в Node.js Foundation, координируя маркетинговую стратегию и реализацию, включая поддержку слияния.
Трейси Ли (GitHub: @ ladyleet, [email protected], Twitter: @ladyleet) продолжает служить нашим связующим звеном с сообществом Node.js, занимаясь логистикой мероприятий сообщества, координацией дорожных фондов и вопросами, связанными с хабаром.
Если у вас есть вопросы, не стесняйтесь обращаться к нам! Мы очень рады тому, что нас ждет в 2019 году и чего мы обязательно добьемся вместе.Если у вас возникнут сомнения относительно того, к кому обратиться с вопросом, свяжитесь со всеми нами по электронной почте info@nodejs.