Данная страница не найдена
Сталь мартенситного класса Сложнофазная сталь High Edge Ductility Dual Phase with high formability Двухфазная сталь Сталь, упрочняющаяся при деформации (PHS) Высокопрочная низколегированная сталь (HSLA) Трубы Виды покрытия
По функциональным возможностям По сегментам По названию продукта
Листовая сталь Труба Палец Wire
Предлагаемые варианты поиска
читать, слушать онлайн на Smart Reading
Пластичность. Гибкое мышление в век перемен (Леонард Млодинов) – саммари на книгу: читать, слушать онлайн на Smart Reading
Гибкое мышление в век перемен (Леонард Млодинов) – саммари на книгу: читать, слушать онлайн на Smart ReadingLeonard Mlodinow
Elastic: Flexible Thinking in a Time of Change Leonard Mlodinow 2018
Текст • 22 мин
Аудио • 32 мин
Инфографика • 5 мин
Читать бесплатно 7 дней Попробовать бесплатно 7 дней
О книге
Что общего между Pokémon Go и «Франкенштейном» Мэри Шелли? Почему одни компании выживают, а другие терпят неудачу при первых признаках перемен? Какое неоспоримое преимущество отличает человеческий мозг от компьютера? Ответ: гибкость мышления. От природы все люди обладают способностью справляться с изменениями, но отточенная до совершенства гибкость мышления помогает добиться успехов в работе и повседневной жизни. Леонард Млодинов, например, благодаря ей превратился из профессора физики в знаменитого сценариста. С помощью новейших методов нейробиологии он объясняет принципы, благодаря которым организации адаптируются к новым технологиям, а люди — полностью меняют свою жизнь. Его методика позволяет благоденствовать в мире, где способность к адаптации ценится как никогда прежде.
От природы все люди обладают способностью справляться с изменениями, но отточенная до совершенства гибкость мышления помогает добиться успехов в работе и повседневной жизни. Леонард Млодинов, например, благодаря ей превратился из профессора физики в знаменитого сценариста. С помощью новейших методов нейробиологии он объясняет принципы, благодаря которым организации адаптируются к новым технологиям, а люди — полностью меняют свою жизнь. Его методика позволяет благоденствовать в мире, где способность к адаптации ценится как никогда прежде.Об авторе
Леонард Млодинов — американский писатель. Имеет докторскую степень Калифорнийского университета в Беркли в области теоретической физики. Автор пяти бестселлеров по версии The New York Times. Два из них написаны в соавторстве с физиком Стивеном Хокингом: «Кратчайшая история времени» и «Высший замысел». Читал лекции в Калифорнийском технологическом институте, как аналитик писал статьи для журналов The Wall Street, Forbes. Был одним из сценаристов телесериала «Star Trek: Следующее поколение».
Был одним из сценаристов телесериала «Star Trek: Следующее поколение».
Поделиться в соцсетях
Узнайте, что такое саммариСаммари Smart Reading — краткое изложение ключевых мыслей нехудожественной книги. Главная особенность наших саммари — глубина и содержательность: мы передаем все ценные идеи книги, ее мотивационную составляющую, сохраняем важные примеры, кейсы и даже дополняем текст комментариями, позволяющими глубже понять идеи автора.
Вы прослушали аудиосаммари по книге
«Пластичность. Гибкое мышление в век перемен» автора Леонард Млодинов
Срок вашей подписки истек.
Срок вашей корпоративной подписки истек. Пожалуйста, свяжитесь с отделом продаж [email protected], чтобы оплатить подписку.
Вы успешно подписались на рассылку
Изменить пароль
Это и другие саммари доступны для наших
подписчиков. Попробуйте 7 дней бесплатно или войдите в ваш аккаунт
Попробовать бесплатно
или
Войти в систему
По вопросам корпоративной подписки обращайтесь по адресу
corp@smartreading. ru
ru
Вы уже купили автоматически обновляемую (рекуррентную) подписку. По окончанию срока действия подписки — деньги будут списаны с вашей карты автоматически и подписка будет обновлена.
Вы являетесь корпоративным пользователем. По вопросам продления подписки обращайтесь к Куратору в рамках вашей компании.
У вас уже есть Бессрочная подписка.
У вас уже есть Семейная подписка.
Вы успешно {{ pageTariff_successPayText }} тариф
«{{ pageTariff_PaidTariffName }}»
 Армянский, д. 9 стр.1, офис 309
119021
г. Москва, Телефон:+7 495 260-14-47,
Электронная почта: [email protected]
Армянский, д. 9 стр.1, офис 309
119021
г. Москва, Телефон:+7 495 260-14-47,
Электронная почта: [email protected]14 Экологическая пластичность — СтудИзба
ЛЕКЦИЯ 12
Экологическая пластичность.
Несмотря на большое разнообразие экологических факторов, в характере их воздействия и в ответных реакциях живых организмов можно выявить ряд общих закономерностей. Эффект влияния факторов зависит не только от характера их действия (качества), но и от количественного значения, воспринимаемого организмами, высокая или низкая температура, степень освещенности, влажности, количество пищи и т. д. В процессе эволюции выработалась способность организмов адаптироваться к экологическим факторам в определенных количественных пределах. Уменьшение или увеличение значения фактора за этими пределами угнетает жизнедеятельность, а при достижении некоторого минимального или максимального уровня наступает гибель.
Зоны действия экологического фактора и теоретическая зависимость жизнедеятельности организма популяции или сообщества от количественного значения фактора в общем виде показаны на рис. 5.11.
Количественный диапазон фактора, наиболее благоприятный для жизнедеятельности, называется экологическим оптимумом. Значения фактора, лежащие в зоне угнетения, называются экологическим пессимумом. Минимальное и максимальное значения фактора, при которых наступает гибель, называются соответственно эколгическим минимумом и экологическим максимумом. Кривая изображенная на рис. 5.11 не является симметричной.
Любые виды организмов, популяций или сообществ приспособлены, например, к существованию в определенном интервале температур. Причем температурный диапазон жизни на суше больше, чем в воде. Выносливость водных организмов к ее колебаниям меньше, чем наземных. Верхним пределом жизни, вероятно, являются температуры, при которых разрушаются ферменты и свертываются белки (50 — 60 ОС). Однако отдельные организмы могут существовать при более высоких температурах. Нижний предел температуры, при котором возможна жизнь, около -70 °С. В анабиозе, т. е. в неактивном состоянии, некоторые организмы сохраняются при абсолютном нуле (-273 °С).
Однако отдельные организмы могут существовать при более высоких температурах. Нижний предел температуры, при котором возможна жизнь, около -70 °С. В анабиозе, т. е. в неактивном состоянии, некоторые организмы сохраняются при абсолютном нуле (-273 °С).
Свойства организмов адаптироваться к существованию в том или ином диапазоне экологического фактора называется экологической пластичностью.
Чем шире диапазон экологического фактора, в пределах которого данный организм может жить, тем больше его экологическая пластичность. По степени пластичности выделяют два типа организмов: стенобионтные (стеноэки- узкоприспособленные) и эврибионтные (эвриэки -широкоприспособленные).
Рекомендуемые файлы
Стенобионтные и эврибионтные организмы различаются диапазоном экологического фактора, в котором они могут жить. Стеноэки способны существовать лишь при небольших отклонениях фактора от оптимального значения. Эвриэки -организмы, выдерживающие большую амплитуду колебаний экологического фактора.
Таким образом, стенобионты экологически непластичны, т. е. маловыносливы, а эврибионты экологически пластичны, т. е. более выносливы. К первым относятся, например, типичные обитатели морей, которые живут в условиях высокой солености (камбала), и типичные обитатели пресных вод (карась). Они обладают невысокой экологической пластичностью, в то время как трехиглая колюшка, может жить как в пресных, так и в соленых водах, т. е. характеризуется высокой пластичностью (рис. 5.13).
Количественный диапазон фактора, наиболее благоприятный для жизнедеятельности, называется экологическим оптимумом.
Значения фактора, лежащие в зоне угнетения, называются экологическим пессимумом. Минимальное и максимальное значения фактора, при которых наступает гибель, называются соответственно экологическим минимумом и экологическим максимумом. Организмы, живущие длительное время в относительно стабильных условиях, утрачивают экологическую пластичность, а те, которые были подвержены значительным колебаниям фактора, становятся более выносливыми к нему, т. е. увеличивают экологическую пластичность. Для обозначения отношения организмов к конкретному фактору к его названию прибавляют приставки: стено- и эври-. По отношению к температуре – это стено- или эвритермальные виды. По отношению к солености – стено- или эвригалинные, к свету – стено- или эврифотные и т. д. Стено- или эврибионты проявляются, как правило, по отношению к одному или немногим факторам. Так, эвритермное растение может бытъ стеногигробионтным (невынос к колебаниям влажности), а стеногалинная рыба оказывается эвритермной и т. п. Исторически, приспосабливаясь к экологическим факторам, животные, растения, микроорганизмы распределяются по различным средам, формируя все многообразие экосистем, образующих в итоге биосферу 3емли.
е. увеличивают экологическую пластичность. Для обозначения отношения организмов к конкретному фактору к его названию прибавляют приставки: стено- и эври-. По отношению к температуре – это стено- или эвритермальные виды. По отношению к солености – стено- или эвригалинные, к свету – стено- или эврифотные и т. д. Стено- или эврибионты проявляются, как правило, по отношению к одному или немногим факторам. Так, эвритермное растение может бытъ стеногигробионтным (невынос к колебаниям влажности), а стеногалинная рыба оказывается эвритермной и т. п. Исторически, приспосабливаясь к экологическим факторам, животные, растения, микроорганизмы распределяются по различным средам, формируя все многообразие экосистем, образующих в итоге биосферу 3емли.
Лимитирующие факторы.
Представление о лимитирующих факторах основывается на двух законах экологии: законе минимума и законе толерантности.
Закон минимума. В середине прошлого века немецкий химик Ю. Либих (1840), изучая влияние разнообразных питательных веществ на рост растений, обнаружил, что урожай зависит не от тех элементов питания, которые требуются в больших количествах и присутствуют в изобилии (например, СО2), а от тех, которые, хотя и нужны растению в меньших количествах, но практически отсутствуют в почве или недоступны (например, фосфор, цинк, бор). Эту закономерность Либих сформулировал так: «Рост растения зависит от того элемента питания, который присутствует в минимальном количестве».
Эту закономерность Либих сформулировал так: «Рост растения зависит от того элемента питания, который присутствует в минимальном количестве».
Позднее этот вывод стал известен как закон минимума Либиха и был распространен на другие экологические факторы. Ограничивать, или лимитировать развитие организмов могут тепло, свет, вода, кислород, и другие факторы, если их значение соответствует экологическому минимуму.
Таким образом, закон минимума Либиха можно сформулировать в общем виде так: рост и развитие организмов зависят в первую очередь от тех факторов природной среды, значение которых приближается к экологическому минимуму.
Дальнейшие исследования показали, что закон минимума имеет два ограничения, которые следует у при практическом применении. Первое ограничение состоит в том, что закон Либиха строго применим лишь в условиях стационарного состояния системы. Второе ограничение связано с взаимодействием нескольких факторов. Иногда организм способен заменить (хотя бы частично) дефицитный элемент другим, химически близким.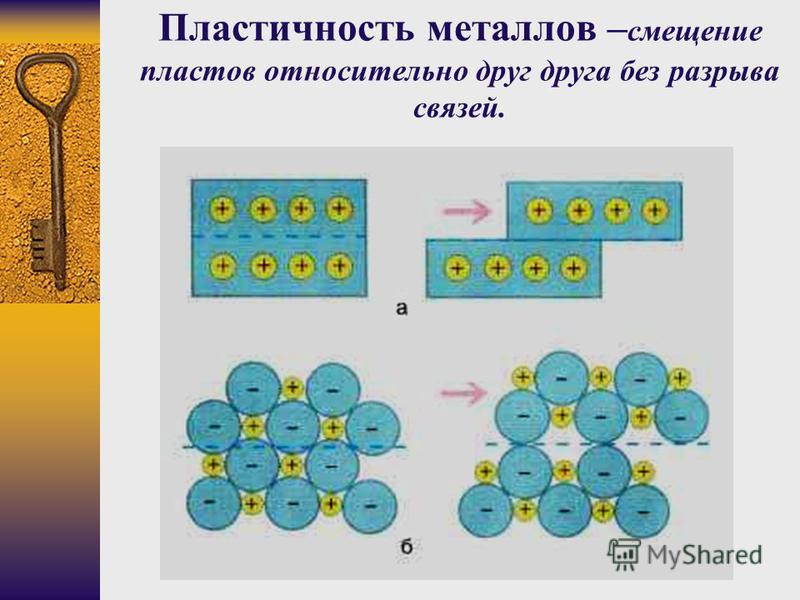
Закон толерантности (ерпения) был открыт английским биологом В. Шелфордом (1913), который обратил внимание на то, что ограничивать развитие живых организмов могут не только те экологические факторы, значения которых минимальны, но и те, которые характеризуются экологическим максимумом. Избыток тепла, света, воды и даже питательных веществ может оказаться столь же губительным, как и их недостаток. Диапазон экологического фактора между минимумом и максимумом В. Шелфорд назвал пределом толерантности. Позднее были проведены многочисленные исследования, которые позволили установить пределы толерантности, т. е. возможного существования, для многих растений и животных. Законы Ю. Либиха и В. Шелфорда помогли понять многие явления и распределение организмов в природе.
Закон толерантности Шелфорда можно сформулировать в общем виде так: рост и развитие организма зависят в первую очередь от тех факторов среды, значения которых приближаются к экологическому минимуму или экологическому максимуму.
Было установлено следующее:
— организмы с широким диапазоном толерантности ко всем факторам широко распространены в природе и часто бывают космополитами. Например, многие патогенные бактерии;
— организмы могут иметь широкий диапазон толерантности в отношении одного фактора и узкий диапазон относительно другого. Например, люди более выносливы к отсутствию пищи, чем к отсутствию воды, т. е предел толерантности относительно воды более узкий, чем относительно пищи;
— если условия по одному из экологических факторов становятся неоптимальными, то может измениться и предел толерантности по другим факторам. Например, при недостатке азота в почве злакам требуется гораздо больше воды;
— наблюдаемые в природе реальные пределы толерантности меньше, чем потенциальные возможности организма адаптированного к данному фактору. Это объясняется тем, что в природе пользоваться оптимальными физическими условиями среды часто мешают биотические отношения (конкуренция, отсутствие опылителей, хищники) и другие взаимодействия факторов.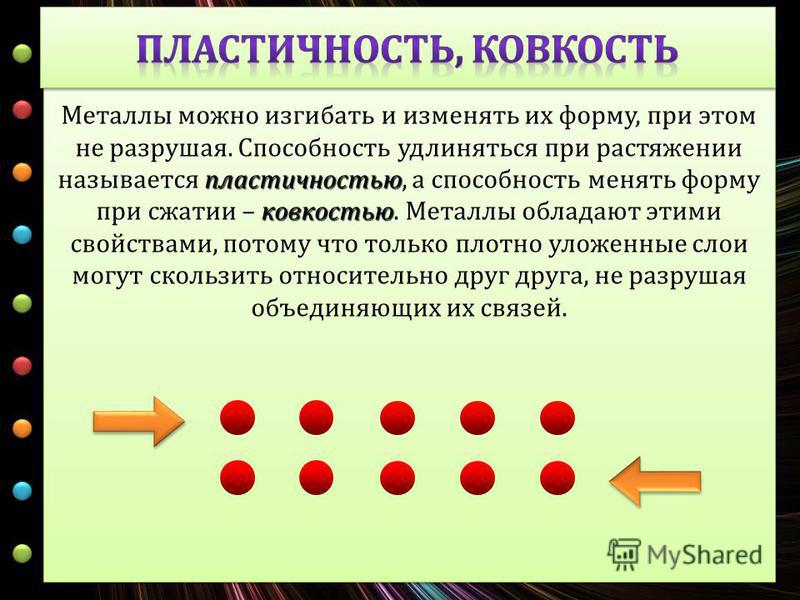 Различают потенциальную и реализованную экологические ниши.
Различают потенциальную и реализованную экологические ниши.
— пределы толерантности у размножающихся особей и потомства меньше, чем у взрослых особей, т. е. самки в период размножения и их потомство менее вы к условиям жизни, чем взрослые организмы. Забота о потомстве и бережное отношение к материнству продиктованы законами природы. К сожалению, иногда социальные достижения противоречат этим законам;
— экстремальные (стрессовые) значения одного из факторов ведут к снижению предела толерантности по другим факторам.
Если значение хотя бы одного из известных факторов приближается к минимуму или максимуму, существование и процветание организма, популяции или сообщества становится зависимым именно от этого лимитирующего жизнедеятельность фактора.
Обобщающая концепция лимитирующих факторов.
Лимитирующим фактором называется любой экологический фактор, приближающийся к крайним значениям пределов толерантности или превышающий их. Такие сильно отклоняющиеся от оптимума факторы приобретают первостепенное значение в жизни организмов и биологических систем, т.к. контролируют условия существования. Наиболее важными лимитирующими факторами на суше являются свет, температура и вода, а море – свет, температура и соленость. Все факторы среды зависят друг от друга и действуют согласованно. Например, содержание кислорода в наземных местообитаниях велико, и он настолько доступен, что практически никогда не служит лимитирующим фактором, следовательно, он мало интересует экологов, занимающихся наземными экосистемами. Напротив, в воде кислород является фактором, лимитирующим развитие живых организмов (рыб). Лимитирующие факторы определяют и географический ареал вида. Так, продвижение организмов на север лимитируется недостатком тепла. Биотические факторы также часто ограничивают распространение тех или иных организмов. Например, завезенный из Средиземноморья в Калифорнию инжир не плодоносил до тех пор, пока не завезли туда определенный вид осы — единственного опылителя этого растения.
Такие сильно отклоняющиеся от оптимума факторы приобретают первостепенное значение в жизни организмов и биологических систем, т.к. контролируют условия существования. Наиболее важными лимитирующими факторами на суше являются свет, температура и вода, а море – свет, температура и соленость. Все факторы среды зависят друг от друга и действуют согласованно. Например, содержание кислорода в наземных местообитаниях велико, и он настолько доступен, что практически никогда не служит лимитирующим фактором, следовательно, он мало интересует экологов, занимающихся наземными экосистемами. Напротив, в воде кислород является фактором, лимитирующим развитие живых организмов (рыб). Лимитирующие факторы определяют и географический ареал вида. Так, продвижение организмов на север лимитируется недостатком тепла. Биотические факторы также часто ограничивают распространение тех или иных организмов. Например, завезенный из Средиземноморья в Калифорнию инжир не плодоносил до тех пор, пока не завезли туда определенный вид осы — единственного опылителя этого растения. Выявление лимитирующих факторов очень важно во многих видах деятельности, особенно в сельском хозяйстве. Если целенаправленно влиять на лимитирующие условия, можно быстро и эффективно повышать урожайность растений и производительность животных. Так, при разведении пшеницы на кислых почвах никакие агрономические мероприятия не дадут эффекта, если не применять известкование, которое снизит ограничивающее действие кислот. Умелое регулирование условиями существования может дать эффективные результаты управления.
Выявление лимитирующих факторов очень важно во многих видах деятельности, особенно в сельском хозяйстве. Если целенаправленно влиять на лимитирующие условия, можно быстро и эффективно повышать урожайность растений и производительность животных. Так, при разведении пшеницы на кислых почвах никакие агрономические мероприятия не дадут эффекта, если не применять известкование, которое снизит ограничивающее действие кислот. Умелое регулирование условиями существования может дать эффективные результаты управления.
Ценность концепции лимитирующих факторов состоит в том, что она позволяет разобраться в сложных взаимосвязях в экосистемах. Приоритетными в тот или иной отрезок времени оказываются различные лимитирующие факторы, которые являются основой при изучении экосистем и управлении ими.
Важным лимитирующим фактором в современных условиях является загрязнение природной среды. Оно происходит в результате внесения в среду веществ, которых в ней не было (металлы, новые синтезированные химические вещества) и которые не разлагаются вовсе, либо существующих в биосфере (например, углекислый газ), но вносимый в чрезмерно больших количествах, не дающих возможности переработать их естественным способом. Образно говоря, загрязняющие вещества — это ресурсы не на своем месте.
Образно говоря, загрязняющие вещества — это ресурсы не на своем месте.
Обратите внимание на лекцию «Инфекционная анемия лошадей».
Отсюда следует, что загрязнение приводит к нежелательному изменению физических, химических и биологических характеристик среды, которое оказывает неблагоприятное влияние на экосистемы и человека. Загрязнение увеличивается как в результате роста населения и его потребностей, так и в результате использования новых технологий, обслуживающих эти потребности. Оно бывает химическим, тепловым, шумовым.
Главный лимитирующий фактор, по Ю. Одуму, — это размеры и качество ойкоса, а не просто число калорий, которые можно выжать из земли. Ландшафт не только склад запасов, но и дом, в котором мы живем. Следует стремиться сохранить по меньшей мере треть всей суши в качестве охраняемого открытого пространства. Это означает, что треть всей нашей среды обитания должны составлять национальные парки, заповедники, зеленые зоны, участки дикой природы. Ограничение использования земли является аналогом природного регулирующего механизма, называемого территориальным поведением. При помощи этого механизма многие виды животных избегают скученности и вызываемого ею стресса.
Ограничение использования земли является аналогом природного регулирующего механизма, называемого территориальным поведением. При помощи этого механизма многие виды животных избегают скученности и вызываемого ею стресса.
Взаимодействие и компенсация факторов. В природе экологические факторы действуют не независимо друг от друга. Анализ влияния одного фактора на организм или сообщество не самоцель, а способ оценки сравнительной значимости различных условий, действующих совместно в реальных экосистемах. Температура и влажность — самые важные климатические факторы в наземных местообитаниях. Взаимодействие этих двух факторов формирует два основных типа климата: морской и континентальный. Водоемы смягчают климат суши, так как вода обладает высокой удельной теплотой плавления и теплоемкостью. Поэтому морскому климату, который формируется вблизи больших озер и морей, свойственны менее резкие колебания и температуры, и влажности, чем континентальному. Воздействие температуры и влажности на организмы также зависит от соотношения их абсолютных значений.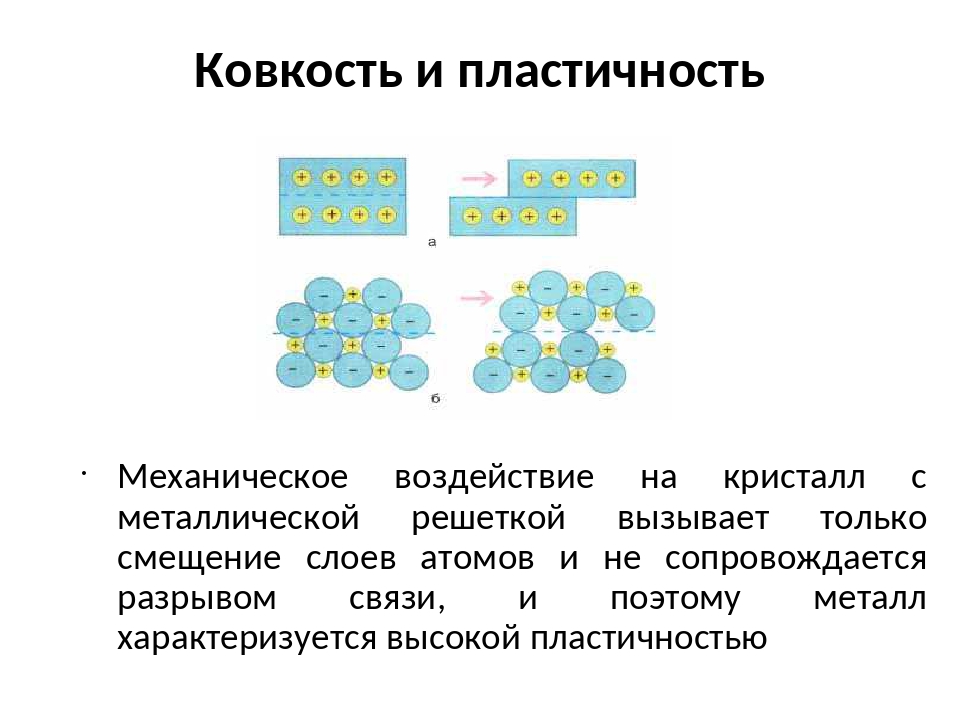 Так, температура оказывает более выраженное лимитирующее влияние, если влажность очень велика или очень мала. Высокие и низкие температуры переносятся хуже при высокой влажности, чем при умеренной. Организмы приспосабливаются к условиям существования и изменяют их, т. е. компенсируют отрицательное воздействие экологических факторов.
Так, температура оказывает более выраженное лимитирующее влияние, если влажность очень велика или очень мала. Высокие и низкие температуры переносятся хуже при высокой влажности, чем при умеренной. Организмы приспосабливаются к условиям существования и изменяют их, т. е. компенсируют отрицательное воздействие экологических факторов.
Компенсация экологических факторов — это стремление организмов ослабить лимитирующее действие физических, биотических и антропогенных влияний. Компенсация факторов возможна на уровне организма, и наиболее эффективна на уровне сообщества. Один и тот же вид, имеющий широкое географическое распространение, может приобретать физиологические и морфологические (гр, форма, очертание) особенности, адаптированные к местным условиям. Например, у животных уши, хвосты, лапы тем короче, а тело тем массивнее, чем холоднее климат. У животных с хорошо развитой моторной активностью компенсация факторов возможна благодаря адаптивному поведению. Возникающие в процессе адаптации изменения часто генетически закрепляются.
Естественную периодичность изменений экологических факторов организмы используют для распределения своих функций во времени. На уровне сообщества компенсация факторов может осуществляться сменой видов по градиенту условий среды. Например, поведение организмов во времени в зависимости от длины дня. Амплитуда длины дня возрастает с географической широтой, что позволяет организмам учитывать не только время года, но и широту местности. Фотопериод — это пусковой механизм последовательности физиологических процессов. Он определяет рост и цветение растений, линьку, миграции и размножение у птиц и млекопитающих и т. д. Фотопериод связан с 6иологическими часами и служит универсальным механизмом регулирования функций во времени. Биологические часы связывают ритмы экологических факторов с физиологическими ритмами, позволяют организмам приспосабливаться к суточной, сезонной, приливно- отливной и другой динамике факторов. Изменяя фотопериод, можно вызывать и желаемые изменения функций организма. . У многих высших организмов адаптация к фотопериоду закрепляется генетически, т. е биологические часы могут работать и при отсутствии закономерной суточной или сезонной динамики.
. У многих высших организмов адаптация к фотопериоду закрепляется генетически, т. е биологические часы могут работать и при отсутствии закономерной суточной или сезонной динамики.
Таким образом, смысл анализа условий среды не в том, чтобы составить перечень экологических факторов, а в том, чтобы обнаружить функционально важные лимитирующие факторы и оценить в какой степени состав и структура экосистем зависит от взаимодействия этих факторов. Только в этом случае удается достоверно прогнозировать результаты изменений и нарушений и управлять экосистемами.
Металлы и неметаллы — урок. Химия, 8–9 класс.
Простые вещества по их свойствам делят на металлы и неметаллы.
Металлы имеют немолекулярное строение и сходные физические свойства. Все металлы (кроме ртути) при обычных условиях представляют собой твёрдые вещества. Их легко узнать по характерному металлическому блеску. Металлы хорошо проводят тепло и электрический ток.
Их легко узнать по характерному металлическому блеску. Металлы хорошо проводят тепло и электрический ток.
Рис. \(1\). Ртуть
Рис. \(2\). Железо
При ударе металлы не разрушаются, а меняют свою форму, т. е. им характерна пластичность. Металлы можно ковать, прокатывать в листы, вытягивать в проволоку.
Неметаллы не имеют общих физических свойств и не похожи на металлы. У них отсутствует металлический блеск. У большинства неметаллов низкие электропроводность и теплопроводность.
Большинство неметаллов имеет молекулярное строение. Такие вещества при обычных условиях являются газами (водород, кислород, азот, озон, фтор, хлор, инертные газы), жидкостями (бром) или хрупкими легкоплавкими твёрдыми веществами (сера, иод, белый фосфор).
Рис. \(3\). Хлор
Рис. \(4\). Бром
Рис. \(5\). Сера
\(5\). Сера
Некоторые неметаллы имеют немолекулярное строение, например, красный фосфор, кремний, алмаз и графит. Такие вещества твёрдые, тугоплавкие, нелетучие.
Рис. \(6\). Уголь
Определить, является простое вещество металлом или неметаллом, можно с помощью периодической таблицы. Химические элементы металлы, образующие простые вещества с металлическими свойствами, располагаются в периодической таблице слева ниже диагонали «водород — бор — кремний — мышьяк — теллур — астат — № \(118\)». Вверху справа располагаются химические элементы неметаллы, которые образуют простые вещества с неметаллическими свойствами.
Рис. \(7\). Периодическая таблица
Элементов металлов больше, чем элементов неметаллов. Значит, и простых веществ с металлическими свойствами существует больше, чем с неметаллическими.
Источники:
Рис. 1. Ртуть https://www.shutterstock.com/ru/image-illustration/shiny-mercury-hg-metal-drops-droplets-373508821
1. Ртуть https://www.shutterstock.com/ru/image-illustration/shiny-mercury-hg-metal-drops-droplets-373508821
Рис. 2. Железо https://www.shutterstock.com/ru/image-photo/99-fine-electrolytic-iron-isolated-on-1687925125
Рис. 3. Хлор https://image.shutterstock.com/image-photo/chlorine-gaz-glass-round-bottom-600w-713676862.jpg
Рис. 4. Бром https://image.shutterstock.com/image-photo/macroview-on-ampoule-element-no-600w-1739647871.jpg
Рис. 5. Сера https://www.shutterstock.com/ru/image-photo/yellow-natural-native-sulfur-crystal-isolated-729259936
Рис. 6. Уголь https://www.shutterstock.com/ru/image-photo/activated-charcoal-isolated-on-white-background-1446133922
Рис. 7. Периодическая таблица © ЯКласс
как мозг меняется в течение жизни и почему возраст — не помеха обучаемости
Человеческий мозг состоит из десятков миллиардов нейронов. По мере взросления они соединяются в масштабную сеть, которая меняется всю жизнь под воздействием внешней среды. Такая пластичность позволяет улучшать когнитивные способности уже во взрослом возрасте, восстанавливаться после серьезных повреждений и решать самые неожиданные задачи.
 T&P публикуют отрывок из книги современного философа Катрин Малабу о том, почему мозг напоминает скульптуру, как определенные занятия помогают улучшить память и есть ли пределы у пластичности.
T&P публикуют отрывок из книги современного философа Катрин Малабу о том, почему мозг напоминает скульптуру, как определенные занятия помогают улучшить память и есть ли пределы у пластичности.1. Пластичность развития: формирование нейронных связей
[…] «Головной мозг человека, — утверждает Жан-Пьер Шанже, — предстает перед нами как гигантская сборка десятков миллиардов спутанных друг с другом нейронных „паутин“, где „стрекочут“ и распространяются тысячи электрических импульсов, тут и там перехватываемых широким спектром химических сигналов». Эти «паутины», также называемые «разветвлениями», — нейронные соединения, постепенно образующиеся в ходе развития индивида. Когда говорят о пластичности, характеризуют именно этот нейрогенез. Мозг, в сущности, формирует сам себя. «Человек рождается с мозгом, который весит 300 граммов, то есть в пять раз меньше мозга взрослого… Одна из главных особенностей развития человеческого мозга состоит в том, что оно продолжается еще длительное время после рождения… примерно пятнадцать лет».
Таким образом, все начинается с установления соединений, затем их умножения и усложнения. Увеличение массы мозга совпадает с разрастанием аксонов и дендритов, образованием синапсов, развитием вокруг аксонов миелиновых оболочек. Это развитие подчинено строгому генетическому детерминизму. По своему происхождению и строению «все человеческие мозги схожи между собой», говорит Марк Жанро. Безусловно, соединения, составляющие анатомию зрелого мозга, не являются делом случая или стихийной организации. Миграция нервных клеток и их приспособление к конечным целям запрограммированы. «Приведем лишь один пример, — продолжает автор. — У всех людей волокна, исходящие из сетчатки и несущие зрительную информацию, завершают свой путь в зрительной части коры больших полушарий, то есть в затылочной доле, находящейся в задней части мозга; у всех людей эта зрительная часть образует связи с другими участками, расположенными в теменной, височной доле и т. д. Следовательно, мозг взрослого человека отражает существование предустановленного плана, который обеспечивает неизменность анатомии от одного индивида к другому».
Если нейрогенез соответствует «предустановленному плану», зачем вообще говорить о пластичности, характеризуя это развитие? По двум существенным причинам, связанным с 1) вышеупомянутым процессом установления соединений и 2) их формовкой (которая отличается от модуляции синаптической эффективности). В обоих случаях выполнение программы действует как пластика. Существует, так сказать, пластическое искусство мозга, чем и объясняется использование термина «пластичность» в данном контексте. Именно здесь на передний план выступает суженное или «закрытое» значение этого понятия — скульптура заранее определенной формы.
С резцом скульптора в процессе установления соединений можно сравнить явление под названием «апоптоз», или «клеточная гибель». Эта смерть представляет собой нормальное явление. Она опять же соответствует выполнению генетической программы, которая приводит к устранению бесполезных связей и постепенному становлению окончательной формы системы за счет нацеливания нервных волокон на клетки-мишени. В человеческом мозге смерть нейронов начинается в конце внутриутробного периода и продолжается как минимум в течение первых шести месяцев жизни. У взрослого человека ее динамика существенно замедляется. Как пишет Жан-Пьер Шанже: «Истребление нейронов — часть нормального развития. Оно составляет один из его решающих этапов». В книге с красноречивым заглавием «Скульптура живого» биолог Жан-Клод Амайзен подчеркивает тот факт, что мозг, отнюдь не будучи, как долго полагалось, полностью сформированным к моменту рождения органом, представляет собой инстанцию, одновременно принимающую и придающую себе собственную форму. «Клеточная гибель, — пишет он, — является… инструментом, позволяющим эмбриону развить свою форму методом исключения, что роднит ее со скульптурой».
В человеческом мозге смерть нейронов начинается в конце внутриутробного периода и продолжается как минимум в течение первых шести месяцев жизни. У взрослого человека ее динамика существенно замедляется. Как пишет Жан-Пьер Шанже: «Истребление нейронов — часть нормального развития. Оно составляет один из его решающих этапов». В книге с красноречивым заглавием «Скульптура живого» биолог Жан-Клод Амайзен подчеркивает тот факт, что мозг, отнюдь не будучи, как долго полагалось, полностью сформированным к моменту рождения органом, представляет собой инстанцию, одновременно принимающую и придающую себе собственную форму. «Клеточная гибель, — пишет он, — является… инструментом, позволяющим эмбриону развить свою форму методом исключения, что роднит ее со скульптурой».
Между тем, как объясняет Марк Жанро, начиная с этой стадии развития и как только система обретает законченную скульптурную форму, «генетический детерминизм ослабевает». «После рождения топографическая сеть, образовавшаяся в ходе эмбриогенеза и стабилизировавшаяся через гибель нейронов и устранение соединений, начинает функционировать под влиянием внешних факторов. Это функционирование влечет за собой новую фазу формовки соединений». Среда играет здесь основополагающую роль.
Это функционирование влечет за собой новую фазу формовки соединений». Среда играет здесь основополагающую роль.
Развитие человеческого мозга по большей части протекает в открытом пространстве, при соприкосновении с раздражителями мира, которые непосредственно воздействуют на рост и объем соединений.
Например, зрительная система не является целиком функциональной при рождении. Синапсы, что соединяют волокна, исходящие из сетчатки, с нейронами зрительной коры, еще не полностью сформированы. И именно информация, полученная извне, активизирует эти синапсы и содействует их созреванию. В этом ключе и принято говорить о формовке синапсов или механизме синаптической пластичности (как мы видим, по-прежнему связанной с генетической программой) во второй фазе развития.
Процесс образования мозга в течение этих двух фаз — установление соединений и их созревание под воздействием среды — выявляет, таким образом, пластичность в выполнении программы. В обоих случаях мозг предстает как сформированная — постепенно изваянная, стабилизированная, поделенная на разные регионы — и в то же время формообразующая инстанция: мало-помалу, по мере того как увеличивается объем соединений, прорисовывается идентичность индивида. И чем дальше, тем больше эта «первая пластичность» утрачивает свой неумолимый детерминизм. Скульптор постепенно начинает импровизировать. В формовке соединений все большую роль играет наша собственная активность: «Наш мозг, формуемый нашей собственной деятельностью, нашими взаимодействиями с внешним миром, равно как и влиянием, испытываемым нами в ходе воспитания, знает нашу историю и жизненный путь. Из этой близости рождается глубинное тождество функционирования нашего мозга и нашего понимания мира, можно даже сказать, тождество взглядов».
И чем дальше, тем больше эта «первая пластичность» утрачивает свой неумолимый детерминизм. Скульптор постепенно начинает импровизировать. В формовке соединений все большую роль играет наша собственная активность: «Наш мозг, формуемый нашей собственной деятельностью, нашими взаимодействиями с внешним миром, равно как и влиянием, испытываемым нами в ходе воспитания, знает нашу историю и жизненный путь. Из этой близости рождается глубинное тождество функционирования нашего мозга и нашего понимания мира, можно даже сказать, тождество взглядов».
На самом деле первый уровень пластичности тесно связан со вторым, поскольку влияние среды постепенно приходит на смену эпигенетической «лепке» и становится все более выраженным. Суженное или «закрытое» значение пластичности очень скоро наталкивается на ее «открытое» значение — «свободу», в которой удивительным образом сходятся детерминизм и неопределенность. И действительно, мы видим, что морфогенез мозга приводит не к установлению ригидной и окончательно закрепленной структуры, но к образованию того, что можно назвать шаблоном. Он затем оттачивается (вылепляется) в ходе развития и далее — уже в меньшей степени, но с неменьшей эффективностью — на протяжении всей жизни. Нервная деятельность предустановленных цепочек, таким образом, приходит на смену скульптурной активности апоптоза. Отныне роль морфогенных факторов играет окружение мозга как органа (формовка соединений), а затем и его внешнее окружение (синаптическая модуляция под влиянием среды).
Он затем оттачивается (вылепляется) в ходе развития и далее — уже в меньшей степени, но с неменьшей эффективностью — на протяжении всей жизни. Нервная деятельность предустановленных цепочек, таким образом, приходит на смену скульптурной активности апоптоза. Отныне роль морфогенных факторов играет окружение мозга как органа (формовка соединений), а затем и его внешнее окружение (синаптическая модуляция под влиянием среды).
Источник: Svetlana Mokrova / istockphoto.com
2. Пластичность модуляции: мозг и его история
В этом пункте мы сразу же встречаем второе поле действия нейропластичности — модификацию нейронных соединений посредством модуляции синаптической эффективности. Несомненно, именно на этом уровне пластичность проявляет себя с наибольшей яркостью и силой и именно здесь она «раскрывает» свое значение. На самом деле существует своего рода нейронное «творчество», не зависящее ни от чего другого, кроме опыта индивида, его жизни и взаимодействий со средой. Это «творчество» не закреплено исключительно за человеческим мозгом, но характерно уже для наиболее рудиментарных нервных систем.
Данная пластичность, состоящая в придании формы ветвлениям и модуляции синаптической эффективности, была впервые выявлена канадским неврологом Дональдом Олдингом Хеббом❓Дональд Олдинг Хебб (1904–1985) — автор труда «Организация поведения: нейропсихологическая теория» (1949). Термин «пластичность» был впервые предложен выдающимся польским неврологом Ежи (Юрием) Конорским, чье видение синаптического функционирования было довольно близко взглядам Хебба (Konorski J. Conditioned Reflexes and Neuron Organization. Cambridge: Cambridge University Press, 1948; Конорски Ю. Интегративная деятельность мозга. М.: Мир, 1970).. В конце 1930-х годов разнообразные экспериментальные наблюдения заставили его отойти от концепции жесткой локализации цепочек памяти в соответствии с моделью рефлекторной дуги Павлова. По его мысли, следовало скорее говорить о существовании «пластичных синапсов», способных адаптировать свою эффективность передачи. Хебб сформулировал гипотезу нейронных цепочек, способных к самоорганизации, то есть к модификации своих соединений в ходе деятельности, необходимой для восприятия и обучения. Синапс — это первостепенное место, где нервная деятельность может оставить след, способный сместить, изменить или преобразовать себя через повторение прошлых действий.
Синапс — это первостепенное место, где нервная деятельность может оставить след, способный сместить, изменить или преобразовать себя через повторение прошлых действий.
Умение синапсов модулировать свою эффективность и модифицировать интенсивность своих ветвлений под действием опыта осуществляется в двух направлениях. Либо эффективность синапса (его способность передавать сигналы от нейрона к нейрону) увеличивается — это «долговременная потенциация (ДВП или LTP)»; либо она уменьшается — это «долговременная депрессия (ДВД или LTD)». Это видно уже на примере такого животного, как аплизия. Ее центральная нервная система проста и состоит из восьми пар ганглиев, расположенных вокруг пищевода, и большого абдоминального ганглия. Аплизия обладает скудным набором поведенческих стереотипов, среди которых можно выделить ряд защитных маневров вроде втягивания сифона или жабры. Вместе с тем сила ее защитного рефлекса модулируется опытом. Повторение неопасных стимуляций на мантии вызывает уменьшение рефлекса (привыкание), которое выражается в снижении амплитуды отступательных движений. Привыкание сопровождается подавлением синаптической активности и пропорциональным уменьшением количества нейротрансмиттера, выделяемого на уровне сенсорно-двигательного синапса❓Нейротрансмиттеры (ацетилхолин, адреналин и др.) обеспечивают перенос нервного импульса c одной на другую сторону синаптической щели. Здесь на смену электричеству приходит химия (принцип передачи нервного сигнала электрический / химический / электрический)..
Привыкание сопровождается подавлением синаптической активности и пропорциональным уменьшением количества нейротрансмиттера, выделяемого на уровне сенсорно-двигательного синапса❓Нейротрансмиттеры (ацетилхолин, адреналин и др.) обеспечивают перенос нервного импульса c одной на другую сторону синаптической щели. Здесь на смену электричеству приходит химия (принцип передачи нервного сигнала электрический / химический / электрический)..
Явления долговременных потенциации и депрессии еще более отчетливо выражены в процессах приспособления, обучения и памяти у птиц. К примеру, черношапочная гаичка запасает еду в кладовых, которые потом безошибочно находит. Исследователям удалось установить, что размер мозговой зоны (гиппокампа)❓Поскольку гиппокамповая формация и парагиппокампальная извилина играют первостепенную роль в процессе запоминания. Ключ к пластичности мозга и поведения — это способность к обучению и запоминанию. А гиппокамп как раз является зоной, преимущественно отвечающей за эти операции. Его повреждение имеет особенно тяжкие и чаще всего необратимые последствия для когнитивной деятельности и памяти., участвующей в этом процессе, у данной птицы больше, чем у других, не имеющих опыта запасания кормов. Стало быть, виды, практикующие запасание, имеют в сравнении с другими ощутимо более крупный гиппокамп. Это изменение является следствием накопления новых нейронов, уменьшения клеточной гибели (апоптоза) и увеличения соединений между нейронами гиппокампа. Последний, таким образом, демонстрирует удивительную структурную пластичность.
Его повреждение имеет особенно тяжкие и чаще всего необратимые последствия для когнитивной деятельности и памяти., участвующей в этом процессе, у данной птицы больше, чем у других, не имеющих опыта запасания кормов. Стало быть, виды, практикующие запасание, имеют в сравнении с другими ощутимо более крупный гиппокамп. Это изменение является следствием накопления новых нейронов, уменьшения клеточной гибели (апоптоза) и увеличения соединений между нейронами гиппокампа. Последний, таким образом, демонстрирует удивительную структурную пластичность.
Потенциация и депрессия представляют собой не просто синаптические процессы, в течение которых одна или несколько стимуляций вызывают немедленное возбуждение. Это еще и долговременные модификации, способные преобразовать форму (изменения размера определенной зоны мозга, варьирование проницаемости регулярно возбуждаемой зоны) и удалить след, с тем чтобы нанести его заново (лабильность следа памяти). Действительно было замечено, что некоторые нервные сети становятся более продуктивными, когда «подавляют» синапсы, которые были задействованы в задачах, приводивших к ошибкам в двигательном обучении. В человеческом мозге эта особенность проявляется со всей очевидностью в ходе всех процессов обучения. Например, при обучении игре на фортепиано механизм подавления входных сигналов, соответствующих ошибочным движениям («промахам»), делает возможным усвоение правильных движений. В случае потенцированных соединений синапсы увеличивают площадь соприкосновения, их проницаемость возрастает, нервная проводимость ускоряется. И наоборот, малоиспользуемый или «подавляемый» синапс имеет тенденцию становиться менее продуктивным. Нейроны в некотором роде сохраняют импульсы стимуляции. Дело обстоит так, как если бы стабилизация воспоминаний происходила исключительно при условии потенциальной дестабилизации общего пейзажа памяти.
В человеческом мозге эта особенность проявляется со всей очевидностью в ходе всех процессов обучения. Например, при обучении игре на фортепиано механизм подавления входных сигналов, соответствующих ошибочным движениям («промахам»), делает возможным усвоение правильных движений. В случае потенцированных соединений синапсы увеличивают площадь соприкосновения, их проницаемость возрастает, нервная проводимость ускоряется. И наоборот, малоиспользуемый или «подавляемый» синапс имеет тенденцию становиться менее продуктивным. Нейроны в некотором роде сохраняют импульсы стимуляции. Дело обстоит так, как если бы стабилизация воспоминаний происходила исключительно при условии потенциальной дестабилизации общего пейзажа памяти.
Итак, долговременная потенциация структурно связана с долговременной депрессией❓Если бы синапсы, и особенно синапсы гиппокампа, только и делали, что укреплялись под действием ДВП, вскоре все они достигли бы максимальной степени эффективности и, следовательно, кодировка новой информации была бы невозможной. , и эта связь составляет дифференцирующую, а точнее, трансдифференцирующую силу нейропластичности. Если воспользоваться аналогией со становлением стволовых клеток, то можно считать, что благодаря своей пластичности нейронные связи всегда способны сменить различие, воспринять отпечаток или от него избавиться, трансформировать свою программу.
, и эта связь составляет дифференцирующую, а точнее, трансдифференцирующую силу нейропластичности. Если воспользоваться аналогией со становлением стволовых клеток, то можно считать, что благодаря своей пластичности нейронные связи всегда способны сменить различие, воспринять отпечаток или от него избавиться, трансформировать свою программу.
Следовательно,
тот факт, что синапсы демонстрируют усиление или ослабление своей эффективности в зависимости от опыта, позволяет установить, что, хотя анатомия мозга у всех людей одинакова, ни один мозг не идентичен другому по своей истории.
Об этом напрямую свидетельствуют явления обучения и памяти. Повторение и привычка играют существенную роль, а это значит, что у нервной цепочки никогда нет четко фиксированной реакции. Пластичность совмещает роль скульптора с функциями художника и воспитателя свободы и автономии. В некотором смысле допустимо утверждать, что синапсы представляют собой резервы мозга на будущее. Они не закреплены и не служат простыми передатчиками нервной информации, но обладают силой формировать или реформировать саму эту информацию. Марк Жанро пишет: «Эффективность синапсов варьируется в зависимости от потоков информации, которая через них проходит: каждый из нас в детстве и на протяжении всей своей жизни испытывает уникальную конфигурацию влияний внешней среды, которая отражается на форме и работе сетей нашего мозга».
Они не закреплены и не служат простыми передатчиками нервной информации, но обладают силой формировать или реформировать саму эту информацию. Марк Жанро пишет: «Эффективность синапсов варьируется в зависимости от потоков информации, которая через них проходит: каждый из нас в детстве и на протяжении всей своей жизни испытывает уникальную конфигурацию влияний внешней среды, которая отражается на форме и работе сетей нашего мозга».
Это вновь позволяет поставить под сомнение старую догму, согласно которой взрослый мозг, как правило, утрачивает свою пластичность — он, конечно, в состоянии накапливать новую информацию, но с его обучаемостью, функциями памяти и глобальной структурой не происходит никаких изменений, разве что в сторону упадка или дегенерации. Мы, напротив, видим, что имеет место постоянная перестройка нейронной морфологии.
3. Восстановительная пластичность: мозг и его регенерация
Здесь мы подходим к рассмотрению третьего поля действия пластичности — восстановления. За термином «восстановительная пластичность» на самом деле кроются два различных процесса: обновление нейронов, или вторичный нейрогенез, и способность мозга восполнять некоторые виды ущерба, вызванного повреждениями.
За термином «восстановительная пластичность» на самом деле кроются два различных процесса: обновление нейронов, или вторичный нейрогенез, и способность мозга восполнять некоторые виды ущерба, вызванного повреждениями.
Что подразумевается под «обновлением нейронов» или «вторичным нейрогенезом»? Из того, что мы только что сказали, вытекает, будто за первой пластичностью — морфогенетической — следует пластичность модуляции, которая модифицирует синаптическую эффективность, но не затрагивает анатомическую стабильность мозга, как если бы она работала, так сказать, внутри закрытой системы. По словам Хезер Кэмерон: «Некоторые ученые до сих пор придерживаются гипотезы стабильного мозга, согласно которой никакой анатомической пластичности взрослого мозга, и особенно коры больших полушарий, не существует; они считают, что функциональная пластичность, лежащая в основе механизмов обучения, предполагает изменение „мощности“ синапсов, производимое модификацией рецепторов или межклеточной среды нейронов на молекулярном уровне»❓Хезер Кэмерон — исследовательница в лаборатории молекулярной биологии Национального института неврологических расстройств и инсульта (NINDS/NIH) в Бетесде, США. Мы цитируем ее статью: Cameron H. Naissance des neurones et mort d’un dogme // La Recherche, No 329, Mars 2000. P. 35.. Однако эта догма стабильного мозга не вполне верна. Действительно, продолжает автор,
Мы цитируем ее статью: Cameron H. Naissance des neurones et mort d’un dogme // La Recherche, No 329, Mars 2000. P. 35.. Однако эта догма стабильного мозга не вполне верна. Действительно, продолжает автор,
«сегодня нам известно, что отдельные нейроны в зонах, важных для процесса обучения, постоянно обновляются — и это составляет относительно существенные анатомические изменения».
Даже если роль стволовых клеток во взрослом мозге и их локализация все еще мало изучены, даже если существует вероятность, что вторичный нейрогенез затрагивает не все регионы мозга, неоспорим тот факт, что нервные клетки во взрослом возрасте обновляются. Это открывает небывалые перспективы для восстановления мозга и меняет наши представления о его функционировании.
Недавнее исследование неокортекса приматов выявило наличие новых нейронов в трех регионах ассоциативной коры: префронтальной, нижней височной и теменной зонах. «Данный результат особенно интересен, поскольку ассоциативная кора играет важную роль в высокоуровневых когнитивных функциях, тогда как стриарная кора [в которой не наблюдается обновления] участвует в обработке информации зрительного происхождения. Это различие наводит на мысль, что для пластичных по своей природе функций нейрогенез играет ключевую роль и в то же время он бесполезен для функций низшего уровня, таких как обработка данных органов чувств, которые, как правило, стабильны в течение всей жизни».
Это различие наводит на мысль, что для пластичных по своей природе функций нейрогенез играет ключевую роль и в то же время он бесполезен для функций низшего уровня, таких как обработка данных органов чувств, которые, как правило, стабильны в течение всей жизни».
Производство новых нейронов, таким образом, имеет целью не просто замену отмирающих клеток. Оно играет роль в пластичности модуляции и в силу этого еще больше расширяет понятие пластичности — вплоть до того, что колеблет понятие стабильности. Опять же: статуя оживает, программа одушевляется. Там, где, как нам кажется, присутствует лишь чистая механика, мы обнаруживаем сложное переплетение разных типов пластичности, противоречащих привычным представлениям о мозге как машине. Как утверждает Ален Прошьянц: «Необходимо сказать, что одна из главных характеристик нервной системы, безусловно, заключается в ее пластичности. Мозг нельзя рассматривать как сеть окончательно проложенных кабелей, а старение мозга — как выключение все возрастающего числа элементов этой цепочки из сети. Даже несмотря на то что это формально доказано лишь в нескольких экспериментальных моделях, мы вправе предположить, что нервные волокна вырастают ежедневно, что одни синапсы распадаются, а другие, новые, образуются. Эти перемены в нейронном пейзаже… являются свидетельством нашей приспособляемости, обучаемости и нашего потенциала к совершенствованию, который сохраняется вплоть до преклонного возраста, а фактически — до самой смерти».
Даже несмотря на то что это формально доказано лишь в нескольких экспериментальных моделях, мы вправе предположить, что нервные волокна вырастают ежедневно, что одни синапсы распадаются, а другие, новые, образуются. Эти перемены в нейронном пейзаже… являются свидетельством нашей приспособляемости, обучаемости и нашего потенциала к совершенствованию, который сохраняется вплоть до преклонного возраста, а фактически — до самой смерти».
В статье под названием «Необычное распределение новых нейронов» исследователи утверждают: «…наблюдения вторичного нейрогенеза отчетливо показывают, что адаптивные способности нервной системы птиц и взрослых млекопитающих проистекают не только из изменчивости синаптических соединений. Они также опираются на производство или обновление отдельных популяций нейронов в нескольких строго определенных регионах, общим свойством которых является выполнение функций, отвечающих за обучение и/или память. В этом свете представляется, что вторичный нейрогенез равным образом позволяет личному опыту субъекта накладывать свой отпечаток на нейронные сети в виде регулярных морфологических и функциональных перестроек. А значит, взрослый нейрогенез в качестве предельного механизма пластичности, в значительной мере управляемого личным опытом субъекта и его взаимодействиями со средой, по всей видимости, составляет дополнительный механизм индивидуации. С той существенной разницей, что действует он в течение всей жизни».
А значит, взрослый нейрогенез в качестве предельного механизма пластичности, в значительной мере управляемого личным опытом субъекта и его взаимодействиями со средой, по всей видимости, составляет дополнительный механизм индивидуации. С той существенной разницей, что действует он в течение всей жизни».
Идея клеточного обновления, регенерации, ресурса как вспомогательных средств синаптической пластичности проливает свет на силу исцеления — лечения, заживления, компенсации, восстановления, способности мозга создавать естественные «протезы». Пластическое искусство мозга рождает статую, способную излечивать саму себя. Функционирование мозга, как мы знаем, может быть нарушено по причине многих патологий, наиболее известные из которых — черепно-мозговые травмы, нарушения мозгового кровообращения, энцефалиты, нейродегенеративные заболевания (болезнь Паркинсона, болезнь Альцгеймера). Вместе с тем после подобных повреждений и недугов нервная система неизменно проявляет пластичность независимо от того, венчаются ее усилия успехом или нет: пораженные структуры и функции пытаются себя модифицировать, с тем чтобы восполнить дефицит или сформировать новую схему организации, одновременно отклоняющуюся от нормы и ее восстанавливающую.
Восстановительная пластичность, очевидно, восполняет не все дефициты. Некоторые поражения, как мы знаем, необратимы. Но поначалу мозг всегда пытается с большей или меньшей результативностью, эффективностью и надежностью реорганизовать пораженную функцию. В качестве примера данного феномена Марк Жанро приводит «паралич левой руки, вызванный поражением двигательной коры справа вследствие инсульта. Сначала никакое движение невозможно, рука неподвижна и вяла. Через некоторое время мышечная сила возвращается, восстанавливается подвижность локтя и запястья. Как это возможно, если нейроны, отвечающие за управление этими движениями, разрушены?… Тут на помощь приходит функциональная нейровизуализация: она показывает, что при усилиях пациента двигать парализованной рукой активизируется как раз не затронутая поражением двигательная кора слева. Пациент, сам по себе или за счет реабилитации, учится использовать нервные пути, которые в нормальном состоянии таковыми не являются. Подобная реорганизация двигательной функции еще раз свидетельствует о пластичности церебральных механизмов». Другой пример — это то, что происходит на начальном этапе болезни Альцгеймера. Наступающая амнезия частично компенсируется способностью восстановления хранящейся информации. Деактивация одних регионов (гиппокампа) уравновешивается метаболической активацией других (фронтальных). Таким образом,
Другой пример — это то, что происходит на начальном этапе болезни Альцгеймера. Наступающая амнезия частично компенсируется способностью восстановления хранящейся информации. Деактивация одних регионов (гиппокампа) уравновешивается метаболической активацией других (фронтальных). Таким образом,
за поражением отдельных цепочек следует смена стратегий обработки информации, которая также свидетельствует о функциональной пластичности мозга.
Итак, в мозге человека предусмотрены функции, ответственные за реорганизацию после повреждений. Эти явления также наблюдаются в случае некоторых трансплантаций. В январе 2000 года сотрудники лионской больницы имени Эдуара Эррио впервые провели пересадку кистей рук тридцатитрехлетнему Дени Шателье, который за три года до этого перенес ампутацию после случайного взрыва. Вопрос стоял следующим образом: даже если удастся установить анатомическую преемственность между кистями донора и предплечьями реципиента, можно ли добиться той же преемственности на психологическом и неврологическом уровнях? Случай Д. Ш. доказал, что да. Фантомные боли у него исчезли, а двигательный прогресс, которого он добился, позволил заключить, что его мозг успешно интегрировал пересаженные руки. «Когда двигательная кора реорганизуется, синапсы модифицируются. Они изменяют свои влияние и „вес“ в локальном функционировании сети нейронов… После пересадки [подобное] изменение нейронных связей могло бы привести к восстановлению представительства кисти руки». Очередное доказательство удивительной приспособляемости нашего мозга.
Ш. доказал, что да. Фантомные боли у него исчезли, а двигательный прогресс, которого он добился, позволил заключить, что его мозг успешно интегрировал пересаженные руки. «Когда двигательная кора реорганизуется, синапсы модифицируются. Они изменяют свои влияние и „вес“ в локальном функционировании сети нейронов… После пересадки [подобное] изменение нейронных связей могло бы привести к восстановлению представительства кисти руки». Очередное доказательство удивительной приспособляемости нашего мозга.
В рубрике «Открытое чтение» мы публикуем отрывки из книг в том виде, в котором их предоставляют издатели. Незначительные сокращения обозначены многоточием в квадратных скобках.
Мнение автора может не совпадать с мнением редакции.
Где можно учиться по теме #здоровье
Где можно учиться по теме #мозг
мозговых центров APEX | Эшвилл Северная Каролина | 3 Реальные примеры пластичности мозга
3 Реальные примеры пластичности мозга. Пластичность мозга, также известная как нейропластичность, представляет собой способность мозга меняться и расти с течением времени в ответ на окружающую среду. Изменения могут происходить быстро или медленно, и они могут быть положительными или отрицательными. Следующие примеры показывают, как Brain Training опирается на нейропластичность, чтобы помочь людям, которые нуждаются в позитивных изменениях. Вы МОЖЕТЕ научить старую собаку новым трюкам!
Изменения могут происходить быстро или медленно, и они могут быть положительными или отрицательными. Следующие примеры показывают, как Brain Training опирается на нейропластичность, чтобы помочь людям, которые нуждаются в позитивных изменениях. Вы МОЖЕТЕ научить старую собаку новым трюкам!
3 реальных примера восстановления после черепно-мозговой травмы
Мозг BL получил ударную волну от взрыва взрывного устройства (СВУ) во время службы нашей стране на Ближнем Востоке.Позже он получил черепно-мозговую травму (ЧМТ), когда его сбила машина. Эти травмы вызвали у БЛ серьезную когнитивную и поведенческую дисфункцию, в первую очередь проблемы гнева и ярости. Эти проблемы значительно ухудшили его способность реинтегрироваться в общество и его профессиональную гражданскую карьеру. Он также страдал от состояния, называемого дисавтономией, в результате травм головы. Это состояние заставляло его ежедневно потреблять невероятное количество калорий (почти 10 тысяч) только для того, чтобы поддерживать свой вес. Благодаря курсу интенсивной тренировки мозга, подкрепленному уходом на дому и лечебным питанием, Б.Л. в настоящее время успешно работает в качестве лицензированного профессионала и продвигается по карьерной лестнице. Его ярость находится под контролем и больше не приводит к опасным ситуациям. Он также может придерживаться обычной диеты (не более 3500 калорий в день), не теряя нездорового веса.
Благодаря курсу интенсивной тренировки мозга, подкрепленному уходом на дому и лечебным питанием, Б.Л. в настоящее время успешно работает в качестве лицензированного профессионала и продвигается по карьерной лестнице. Его ярость находится под контролем и больше не приводит к опасным ситуациям. Он также может придерживаться обычной диеты (не более 3500 калорий в день), не теряя нездорового веса.
СДВГ, СДВГ, ОКР и пластичность мозга
PJ — молодой человек, который всю свою жизнь боролся с проблемами обучения и поведения.Злоупотребление наркотиками и алкоголем усугубило его проблемы и преждевременно отвлекло его от продуктивной жизни. Он бросил среднюю школу незадолго до выпуска и обнаружил, что не может сдать экзамены, эквивалентные средней школе (GED). Вместо того, чтобы работать или ходить в школу, он проводил время в программах реабилитации от наркомании и алкоголизма. Во время программы обучения мозга PJs с APEX он набрал предварительные баллы, которые позволили ему продолжить сдачу всех частей его GED-тестирования, и он получил высокие проходные баллы по всем предметам. С тех пор он поступил в наш местный общественный колледж и уверенно движется к продуктивной жизни. Теперь его мозг способен сосредотачиваться и решать жизненные проблемы!
С тех пор он поступил в наш местный общественный колледж и уверенно движется к продуктивной жизни. Теперь его мозг способен сосредотачиваться и решать жизненные проблемы!
Повышение когнитивных способностей
Б.А. — молодая женщина, которая годами страдала от пренебрежения и жестокого обращения в детстве. Жестокое обращение привело к когнитивной дисфункции и затруднениям в обучении, которые помешали ей научиться читать. Нарушения развития Б.А. вызвали у нее, как и у ее приемной семьи, много эмоциональных потрясений.Ее неспособность «все обдумать» привела к более деструктивному поведению, в том числе к незаконным действиям. Менее чем через неделю после 5-дневной программы интенсивной тренировки мозга BA впервые за почти 5 лет смог взять в руки книгу и прочитать ее от корки до корки с легкостью и полным пониманием. Это то, что ее мозг не позволял ей делать раньше! Ее показатели когнитивного тестирования увеличились, включая улучшения, превышающие 100%, особенно в областях памяти и внимания. Кроме того, теперь она впервые проявляет признаки раскаяния за многие из своих прошлых проступков.Хотя BA далека от полного выздоровления, она уже на пути к тому, чтобы восстановить контроль над своей жизнью и стать продуктивным и полезным членом своего сообщества — 3 реальных примера пластичности мозга!
Кроме того, теперь она впервые проявляет признаки раскаяния за многие из своих прошлых проступков.Хотя BA далека от полного выздоровления, она уже на пути к тому, чтобы восстановить контроль над своей жизнью и стать продуктивным и полезным членом своего сообщества — 3 реальных примера пластичности мозга!
границ | Структурная пластичность: механизмы и вклад в развитие психических расстройств
Введение
Дендритные шипики являются основным местом передачи возбуждения в головном мозге. Обычно они контактируют с проходящими пресинаптическими окончаниями и чаще всего окружены астроцитарными отростками, образуя сложные структуры, демонстрирующие высокую степень функциональной и структурной пластичности.В то время как основное внимание исследователей обычно сосредотачивалось на функциональных аспектах синаптической пластичности и их ключевом вкладе в механизмы обучения и памяти, работа последнего десятилетия ясно продемонстрировала важность связанных структурных перестроек. Они состоят из различных типов морфологических изменений (увеличение, рост, обрезка, стабилизация), влияющих на разных партнеров (шипы, терминальные отростки, астроцитарные отростки) и происходящих в разных временных масштабах (от минут до дней), что иногда затрудняет их соотнесение с функциональные изменения.Эти структурные перестройки также строго контролируются активностью, они обычно зависят от рецептора NMDA и могут значительно влиять на развитие и организацию локальных синаптических сетей. Недавние достижения позволили раскрыть некоторые сложные молекулярные механизмы и сигнальные системы, регулирующие эти синаптические перестройки, особенно на постсинаптическом уровне. Поэтому в этом обзоре мы сосредоточимся в основном на этих аспектах и выделим множество механизмов, которые могут влиять на структурную пластичность и развитие синаптических сетей и тем самым способствовать когнитивным расстройствам.
Они состоят из различных типов морфологических изменений (увеличение, рост, обрезка, стабилизация), влияющих на разных партнеров (шипы, терминальные отростки, астроцитарные отростки) и происходящих в разных временных масштабах (от минут до дней), что иногда затрудняет их соотнесение с функциональные изменения.Эти структурные перестройки также строго контролируются активностью, они обычно зависят от рецептора NMDA и могут значительно влиять на развитие и организацию локальных синаптических сетей. Недавние достижения позволили раскрыть некоторые сложные молекулярные механизмы и сигнальные системы, регулирующие эти синаптические перестройки, особенно на постсинаптическом уровне. Поэтому в этом обзоре мы сосредоточимся в основном на этих аспектах и выделим множество механизмов, которые могут влиять на структурную пластичность и развитие синаптических сетей и тем самым способствовать когнитивным расстройствам.
Морфологическая изменчивость возбуждающих синапсов
Особенностью дендритных шипиков является высокая изменчивость их морфологической организации. Они демонстрируют большие различия в объеме, причем большие шипы в несколько сотен раз больше, чем маленькие шипы, а также по длине, форме и содержанию в органеллах, таких как рибосомы, эндосомальные системы или аппарат шипов. Считается, что эта высокая морфологическая изменчивость отражает различные функциональные свойства возбуждающих синапсов, связанные с размером постсинаптической плотности, количеством постсинаптических рецепторов, вставленных в постсинаптическую плотность, силой синапса, стадией его развития или даже его стабильностью во времени.Таким образом, часто считается, что морфология позвоночника коррелирует с функциональными параметрами (Bourne and Harris, 2008). Большие шипы, обычно называемые шипами грибовидного типа, а иногда также шипами памяти, связаны со зрелыми, стабильными синапсами, которые были усилены в процессе расширения, опосредованного активностью или пластичностью. Напротив, тонкие удлиненные шипы с маленькими головками, иногда называемые обучающимися шипиками, интерпретируются как представляющие собой молодые, недавно сформированные синаптические структуры, которые с большей вероятностью со временем исчезнут (Bourne and Harris, 2007).
Они демонстрируют большие различия в объеме, причем большие шипы в несколько сотен раз больше, чем маленькие шипы, а также по длине, форме и содержанию в органеллах, таких как рибосомы, эндосомальные системы или аппарат шипов. Считается, что эта высокая морфологическая изменчивость отражает различные функциональные свойства возбуждающих синапсов, связанные с размером постсинаптической плотности, количеством постсинаптических рецепторов, вставленных в постсинаптическую плотность, силой синапса, стадией его развития или даже его стабильностью во времени.Таким образом, часто считается, что морфология позвоночника коррелирует с функциональными параметрами (Bourne and Harris, 2008). Большие шипы, обычно называемые шипами грибовидного типа, а иногда также шипами памяти, связаны со зрелыми, стабильными синапсами, которые были усилены в процессе расширения, опосредованного активностью или пластичностью. Напротив, тонкие удлиненные шипы с маленькими головками, иногда называемые обучающимися шипиками, интерпретируются как представляющие собой молодые, недавно сформированные синаптические структуры, которые с большей вероятностью со временем исчезнут (Bourne and Harris, 2007). Несколько исследований in vitro и in vivo показали высокую корреляцию между размером головки позвоночника, размером постсинаптической плотности, размером ответов, вызванных глутаматом, и стабильностью позвоночника (Kasai et al., 2003; Мацудзаки и др., 2004). Помимо тонких шипов, еще один тип выпячиваний, часто анализируемый отдельно, — это филоподии, обычно характеризующиеся отсутствием расширения на кончике. Считается, что филоподии представляют собой предшественников дендритных шипиков (Ziv and Smith, 1996; Petrak et al., 2005; Тони и др., 2007 г.; Kayser et al., 2008), и они в основном наблюдаются на ранних стадиях развития, где они могут составлять до 20% всех протрузий. Однако в тканях подростков и взрослых они составляют лишь несколько процентов всех выпячиваний в зависимости от критериев, используемых для их идентификации, и их функция остается в значительной степени неясной, поскольку большинство филоподий просто появляются и исчезают, не превращаясь в синапсы позвоночника (Zuo et al.
Несколько исследований in vitro и in vivo показали высокую корреляцию между размером головки позвоночника, размером постсинаптической плотности, размером ответов, вызванных глутаматом, и стабильностью позвоночника (Kasai et al., 2003; Мацудзаки и др., 2004). Помимо тонких шипов, еще один тип выпячиваний, часто анализируемый отдельно, — это филоподии, обычно характеризующиеся отсутствием расширения на кончике. Считается, что филоподии представляют собой предшественников дендритных шипиков (Ziv and Smith, 1996; Petrak et al., 2005; Тони и др., 2007 г.; Kayser et al., 2008), и они в основном наблюдаются на ранних стадиях развития, где они могут составлять до 20% всех протрузий. Однако в тканях подростков и взрослых они составляют лишь несколько процентов всех выпячиваний в зависимости от критериев, используемых для их идентификации, и их функция остается в значительной степени неясной, поскольку большинство филоподий просто появляются и исчезают, не превращаясь в синапсы позвоночника (Zuo et al. , 2005). ; Де Ру и др., 2008а). В целом, исходя из нашего современного понимания свойств шипиков, кажется вероятным, что высокая морфологическая изменчивость дендритных шипиков отражает различные стадии созревания возбуждающих синапсов и их индивидуальную историю.Соответственно, изменения морфологии или плотности позвоночника, наблюдаемые при некоторых психических расстройствах, связанных с развитием, часто интерпретируются как указывающие на дефекты морфогенеза, стабильности или пластичности позвоночника. Однако более глубокий анализ динамических свойств дендритных шипов может обеспечить лучшее понимание основных дефектов.
, 2005). ; Де Ру и др., 2008а). В целом, исходя из нашего современного понимания свойств шипиков, кажется вероятным, что высокая морфологическая изменчивость дендритных шипиков отражает различные стадии созревания возбуждающих синапсов и их индивидуальную историю.Соответственно, изменения морфологии или плотности позвоночника, наблюдаемые при некоторых психических расстройствах, связанных с развитием, часто интерпретируются как указывающие на дефекты морфогенеза, стабильности или пластичности позвоночника. Однако более глубокий анализ динамических свойств дендритных шипов может обеспечить лучшее понимание основных дефектов.
Механизм, способствующий структурной пластичности
Крупный прорыв, ставший возможным благодаря разработке подходов к конфокальной визуализации in vivo , заключался в демонстрации того, что дендритные шипики являются высокопластичными структурами, которые не только постоянно меняют форму с течением времени (Matus, 2000; Yasumatsu et al., 2008), но также могут образовываться и элиминироваться в течение жизни в зависимости от активности (Lendvai et al. , 2000; Holtmaat et al., 2005; Caroni et al., 2012). Эти наблюдения поддерживают концепцию динамической регуляции возбуждающих синаптических сетей механизмами активности и пластичности. Поразительным аспектом этой динамической регуляции связности является ее величина в периоды интенсивного развития, такие как критические периоды, и тот факт, что затем она уменьшается с возрастом (Zuo et al., 2005; Holtmaat and Svoboda, 2009).Это согласуется с представлением о том, что контроль динамики позвоночника с помощью активности играет центральную роль в формировании организации локальных синаптических сетей во время развития, тогда как сниженная структурная пластичность, все еще присутствующая у старых животных, поддерживает возможности адаптации, сводя к минимуму возможное нарушение существующих. схемы.
, 2000; Holtmaat et al., 2005; Caroni et al., 2012). Эти наблюдения поддерживают концепцию динамической регуляции возбуждающих синаптических сетей механизмами активности и пластичности. Поразительным аспектом этой динамической регуляции связности является ее величина в периоды интенсивного развития, такие как критические периоды, и тот факт, что затем она уменьшается с возрастом (Zuo et al., 2005; Holtmaat and Svoboda, 2009).Это согласуется с представлением о том, что контроль динамики позвоночника с помощью активности играет центральную роль в формировании организации локальных синаптических сетей во время развития, тогда как сниженная структурная пластичность, все еще присутствующая у старых животных, поддерживает возможности адаптации, сводя к минимуму возможное нарушение существующих. схемы.
Эти синаптические перестройки включают несколько механизмов для контроля образования, поддержания и устранения возбуждающих синапсов.Весьма вероятно, что различные молекулярные пути независимо регулируют эти разные механизмы, поскольку специфические манипуляции могут избирательно влиять на формирование, стабилизацию или устранение протрузий (Mendez et al. , 2010b; Koleske, 2013; Kehoe et al., 2014). Кроме того, вероятно, существуют гомеостатические регуляции, координирующие эти процессы и способные компенсировать изменения того или иного механизма. Например, мы наблюдали, что трансфекция пирамидных нейронов доминантно-негативным мутантом N-кадгерина, который сильно снижает стабильность шипов, также приводит к компенсаторному увеличению образования шипов (Mendez et al., 2010а). Наконец, эти различные механизмы точно настраиваются за счет индукции активности и пластичности, что еще больше усложняет их регуляцию.
, 2010b; Koleske, 2013; Kehoe et al., 2014). Кроме того, вероятно, существуют гомеостатические регуляции, координирующие эти процессы и способные компенсировать изменения того или иного механизма. Например, мы наблюдали, что трансфекция пирамидных нейронов доминантно-негативным мутантом N-кадгерина, который сильно снижает стабильность шипов, также приводит к компенсаторному увеличению образования шипов (Mendez et al., 2010а). Наконец, эти различные механизмы точно настраиваются за счет индукции активности и пластичности, что еще больше усложняет их регуляцию.
В контексте парадигм, связанных с обучением (LTP), по-видимому, существует два основных структурных типа изменений, о которых сообщалось как в системах in vitro , таких как культура органотипических срезов гиппокампа, так и в условиях in vivo у анестезированных мышей. (De Roo et al., 2008b; Hübener and Bonhoeffer, 2010). В культурах срезов индукция долговременной потенциации приводит к двум основным изменениям: увеличению оборота шипов, характеризующемуся увеличением роста шипов, а также элиминацией шипов (Engert and Bonhoeffer, 1999; Nägerl et al. , 2004; De Roo et al., 2008b) и избирательной стабилизации активированных синапсов (De Roo et al., 2008b; Hill, Zito, 2013). У живых мышей аналогичные изменения также наблюдались в зрительной или моторной коре (Keck et al., 2008; Xu et al., 2009). Примечательно, что у мышей, обученных двигательным задачам на ловкость, оборот позвоночника увеличивается и коррелирует со способностью к обучению задаче, в то время как также наблюдается специфическая стабилизация вновь образованных позвоночников, которая коррелирует с памятью или сохранением задачи (Xu et al., 2009; Ян и др., 2009). Основываясь на результатах этого типа, можно предположить, что изменения оборота позвоночника и изменения стабильности позвоночника представляют собой два ключевых механизма, связанных с памятью, которые могут объяснить некоторые из ее антиномических свойств: способность к обучению, которая требует адаптации существующих сетей, и способность сохранять информацию, которая требует поддержания важных функциональных схем (Caroni et al.
, 2004; De Roo et al., 2008b) и избирательной стабилизации активированных синапсов (De Roo et al., 2008b; Hill, Zito, 2013). У живых мышей аналогичные изменения также наблюдались в зрительной или моторной коре (Keck et al., 2008; Xu et al., 2009). Примечательно, что у мышей, обученных двигательным задачам на ловкость, оборот позвоночника увеличивается и коррелирует со способностью к обучению задаче, в то время как также наблюдается специфическая стабилизация вновь образованных позвоночников, которая коррелирует с памятью или сохранением задачи (Xu et al., 2009; Ян и др., 2009). Основываясь на результатах этого типа, можно предположить, что изменения оборота позвоночника и изменения стабильности позвоночника представляют собой два ключевых механизма, связанных с памятью, которые могут объяснить некоторые из ее антиномических свойств: способность к обучению, которая требует адаптации существующих сетей, и способность сохранять информацию, которая требует поддержания важных функциональных схем (Caroni et al.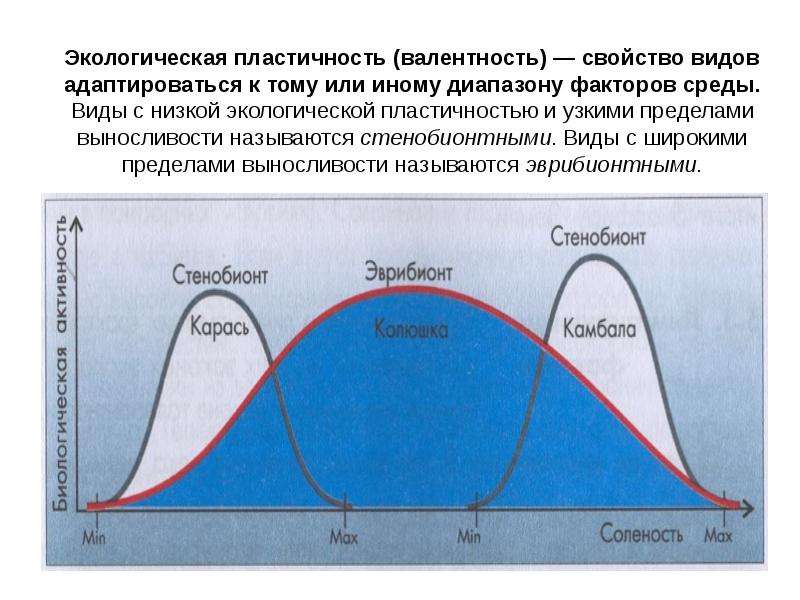 , 2012; см. рис. 1).
, 2012; см. рис. 1).
Рис. 1. Структурная пластичность, обусловленная активностью .Левая панель: синаптические сети характеризуются непрерывным процессом роста (темно-синий шип) и элиминации (пунктирная линия) дендритных шипиков, который регулируется в процессе развития. Средняя панель: во время обучения или деятельности, связанной с обучением, базовый обмен значительно усиливается, что приводит к увеличению образования и ликвидации синаптических контактов, что позволяет ремоделировать и адаптировать связь. Новые шипы также имеют тенденцию расти вблизи активированных синапсов, способствуя образованию кластеров шипов.Правая панель: новообразованные и активированные синапсы предпочтительно стабилизируются (пунктирные кружки), что позволяет поддерживать важные функциональные связи.
Зависящие от активности изменения морфологии и стабильности позвоночника
Идентификация стимулированных синапсов в сети была и остается одним из важных ограничений для понимания того, как активность и пластичность регулируют свойства синапсов. Это было преодолено только в нескольких исследованиях с использованием либо 2-фотонного высвобождения глутамата для достижения локальной стимуляции идентифицированных синапсов, либо визуализации кальция в сочетании с электрической стимуляцией для выявления функциональных синапсов.Хотя предыдущие электронно-микроскопические (ЭМ) анализы уже предполагали, что LTP может быть связана с морфологическими изменениями (Фифкова и Андерсон, 1981; Гейнисман, 1993; Букс и Мюллер, 1996), ранние элегантные исследования группы Касаи (Мацудзаки и др. , 2004) были первыми, кто прямо продемонстрировал, что индукция пластичности действительно связана с быстрым увеличением размера стимулируемых синапсов и повышенной чувствительностью к глутамату. Другие эксперименты также показали, что это увеличение может длиться несколько часов (Zito et al., 2009) и что это коррелирует с реорганизацией актинового цитоскелета (Honkura et al., 2008). Таким образом, эти наблюдения полностью соответствовали накопленным данным, свидетельствующим о причастности актинового цитоскелета и различных актин-регуляторных белков как к поддержанию LTP, так и к морфологии позвоночника (Fukazawa et al.
Это было преодолено только в нескольких исследованиях с использованием либо 2-фотонного высвобождения глутамата для достижения локальной стимуляции идентифицированных синапсов, либо визуализации кальция в сочетании с электрической стимуляцией для выявления функциональных синапсов.Хотя предыдущие электронно-микроскопические (ЭМ) анализы уже предполагали, что LTP может быть связана с морфологическими изменениями (Фифкова и Андерсон, 1981; Гейнисман, 1993; Букс и Мюллер, 1996), ранние элегантные исследования группы Касаи (Мацудзаки и др. , 2004) были первыми, кто прямо продемонстрировал, что индукция пластичности действительно связана с быстрым увеличением размера стимулируемых синапсов и повышенной чувствительностью к глутамату. Другие эксперименты также показали, что это увеличение может длиться несколько часов (Zito et al., 2009) и что это коррелирует с реорганизацией актинового цитоскелета (Honkura et al., 2008). Таким образом, эти наблюдения полностью соответствовали накопленным данным, свидетельствующим о причастности актинового цитоскелета и различных актин-регуляторных белков как к поддержанию LTP, так и к морфологии позвоночника (Fukazawa et al. , 2003; Lisman, 2003; Chen et al., 2007; Bramham). , 2008). Как упоминалось ранее и было обнаружено в исследованиях культур срезов гиппокампа, структурные изменения, влияющие на стимулированные синапсы, также коррелируют со значительным увеличением их долгосрочной стабильности (De Roo et al., 2008б; Хилл и Зито, 2013 г.). Очень немногие стимулированные синапсы исчезали в течение дней после их стимуляции, в то время как нестимулированные синапсы, расположенные на том же дендрите, напротив, имели тенденцию к более быстрой элиминации. Однако остается неясным, связано ли изменение стабильности позвоночника напрямую с увеличением, зависящим от активности. Размер позвоночника и стабильность позвоночника действительно показывают высокую степень корреляции. Повторяющиеся эксперименты с визуализацией показывают, что размер головок позвоночника непрерывно колеблется в течение нескольких дней (Holtmaat et al., 2005; Ясумацу и др., 2008). Кроме того, тот факт, что зависящее от активности увеличение шипов, по-видимому, является временным и полностью исчезает в течение 24 часов, в то время как улучшенная стабильность может быть измерена в течение нескольких дней, предполагает, что они регулируются различными механизмами (De Roo et al.
, 2003; Lisman, 2003; Chen et al., 2007; Bramham). , 2008). Как упоминалось ранее и было обнаружено в исследованиях культур срезов гиппокампа, структурные изменения, влияющие на стимулированные синапсы, также коррелируют со значительным увеличением их долгосрочной стабильности (De Roo et al., 2008б; Хилл и Зито, 2013 г.). Очень немногие стимулированные синапсы исчезали в течение дней после их стимуляции, в то время как нестимулированные синапсы, расположенные на том же дендрите, напротив, имели тенденцию к более быстрой элиминации. Однако остается неясным, связано ли изменение стабильности позвоночника напрямую с увеличением, зависящим от активности. Размер позвоночника и стабильность позвоночника действительно показывают высокую степень корреляции. Повторяющиеся эксперименты с визуализацией показывают, что размер головок позвоночника непрерывно колеблется в течение нескольких дней (Holtmaat et al., 2005; Ясумацу и др., 2008). Кроме того, тот факт, что зависящее от активности увеличение шипов, по-видимому, является временным и полностью исчезает в течение 24 часов, в то время как улучшенная стабильность может быть измерена в течение нескольких дней, предполагает, что они регулируются различными механизмами (De Roo et al. , 2008b).
, 2008b).
Молекулярная природа механизмов, обеспечивающих высокую стабильность активированным шипам, также остается открытым вопросом. Данные различных исследований позволяют предположить, что структурная организация актинового цитоскелета в головке позвоночника может способствовать изменению стабильности позвоночника (Fukazawa et al., 2003; Чен и др., 2007). Фосфорилирование стабилизирующего цитоскелет белка бета-аддуцина, по-видимому, необходимо для стабилизации позвоночника (Bednarek and Caroni, 2011). Кроме того, некоторые молекулы, регулирующие полимеризацию актина, и особенно регуляторы Rho GTPases выше и ниже по течению, влияют на способность шипов увеличиваться и их стабильность (Rex et al., 2009; Murakoshi et al., 2011; Dubos et al., 2012). Белки, которые также могут быть важны для придания стабильности, представляют собой молекулы адгезии, экспрессируемые в возбуждающих синапсах.Предполагается, что нейролигины, интегрины, эфрины и N-кадгерины способствуют сохранению шипиков (Wang et al. , 2008; Matter et al., 2009; Bozdagi et al., 2010; McGeachie et al., 2011; Koleske). , 2013), хотя прямая демонстрация их роли в долговременной стабильности позвоночника, зависящей от активности, часто отсутствует. В исследованиях, проведенных нами для анализа роли N-кадгерина в структурной пластичности, мы обнаружили, что внеклеточный мутант N-кадгерина сильно влияет на продолжительность жизни дендритных шипов и предотвращает стабилизацию шипов, опосредованную активностью (Mendez et al., 2010а). Мы также показали, что N-кадгерин экспрессируется не во всех шипах и что шипы, в которых он экспрессируется, проявляют повышенную стабильность. Кроме того, экспрессия N-кадгерина может быть увеличена за счет паттернов активности, которые индуцируют LTP, и после стимуляции мы обнаружили, что молекула адгезии избирательно накапливается в шипах, которые были активированы. Таким образом, эти результаты предполагают, что изменения в профиле адгезии активированных синапсов способствуют регулированию стабильности и продолжительности жизни синапсов позвоночника (Mendez et al.
, 2008; Matter et al., 2009; Bozdagi et al., 2010; McGeachie et al., 2011; Koleske). , 2013), хотя прямая демонстрация их роли в долговременной стабильности позвоночника, зависящей от активности, часто отсутствует. В исследованиях, проведенных нами для анализа роли N-кадгерина в структурной пластичности, мы обнаружили, что внеклеточный мутант N-кадгерина сильно влияет на продолжительность жизни дендритных шипов и предотвращает стабилизацию шипов, опосредованную активностью (Mendez et al., 2010а). Мы также показали, что N-кадгерин экспрессируется не во всех шипах и что шипы, в которых он экспрессируется, проявляют повышенную стабильность. Кроме того, экспрессия N-кадгерина может быть увеличена за счет паттернов активности, которые индуцируют LTP, и после стимуляции мы обнаружили, что молекула адгезии избирательно накапливается в шипах, которые были активированы. Таким образом, эти результаты предполагают, что изменения в профиле адгезии активированных синапсов способствуют регулированию стабильности и продолжительности жизни синапсов позвоночника (Mendez et al. , 2010а). Вполне вероятно, что этот эффект опосредуется усилением транссинаптических взаимодействий между пре- и постсинаптическими структурами, поскольку в синаптической щели возбуждающих синапсов были идентифицированы молекулярные комплексы, демонстрирующие периодическую организацию, совместимую с существованием N-кадгериновой сети (Zuber et al. ., 2005). Адгезия также может быть усилена между синаптическими структурами и третьим партнером тройного синапса: астроглиальным компонентом. Исследования ЭМ (Лушникова и др., 2009), а также работа Bernardinelli et al.(2014) предполагают, что синаптическая активность и индукция LTP могут управлять реорганизацией астроцитарных процессов вокруг активированных синапсов. Это глиальное структурное ремоделирование опосредовано прямой активацией глиальных метаботропных рецепторов высвобождаемым трансмиттером и притоком кальция в астроцитарный процесс. Это временное увеличение подвижности астроцитарного отростка приводит к лучшему покрытию синапса и повышенной долгосрочной стабильности синаптического контакта.
, 2010а). Вполне вероятно, что этот эффект опосредуется усилением транссинаптических взаимодействий между пре- и постсинаптическими структурами, поскольку в синаптической щели возбуждающих синапсов были идентифицированы молекулярные комплексы, демонстрирующие периодическую организацию, совместимую с существованием N-кадгериновой сети (Zuber et al. ., 2005). Адгезия также может быть усилена между синаптическими структурами и третьим партнером тройного синапса: астроглиальным компонентом. Исследования ЭМ (Лушникова и др., 2009), а также работа Bernardinelli et al.(2014) предполагают, что синаптическая активность и индукция LTP могут управлять реорганизацией астроцитарных процессов вокруг активированных синапсов. Это глиальное структурное ремоделирование опосредовано прямой активацией глиальных метаботропных рецепторов высвобождаемым трансмиттером и притоком кальция в астроцитарный процесс. Это временное увеличение подвижности астроцитарного отростка приводит к лучшему покрытию синапса и повышенной долгосрочной стабильности синаптического контакта. Этот эффект наблюдался как в условиях in vitro , так и в условиях in vivo в соматосенсорной коре и обобщен на рисунке 2.Эти новые результаты, таким образом, указывают на новую роль астроцитарных процессов в зависящей от активности структурной организации синапсов и на вклад астроцитов в генерацию пожизненно устойчивых синапсов. В соответствии с этими наблюдениями недавняя работа также продемонстрировала критическую роль астроцитов в механизмах LTP и долговременной памяти у мышей (Suzuki et al., 2011). И наоборот, существует множество примеров, показывающих, что молекулярное вмешательство в механизмы стабилизации позвоночника, описанные выше, значительно влияет на когнитивные функции (Allen et al., 1998; Судхоф, 2008 г.; Редиес и др., 2012; Вукоевич и др., 2012; Ба и др., 2013).
Этот эффект наблюдался как в условиях in vitro , так и в условиях in vivo в соматосенсорной коре и обобщен на рисунке 2.Эти новые результаты, таким образом, указывают на новую роль астроцитарных процессов в зависящей от активности структурной организации синапсов и на вклад астроцитов в генерацию пожизненно устойчивых синапсов. В соответствии с этими наблюдениями недавняя работа также продемонстрировала критическую роль астроцитов в механизмах LTP и долговременной памяти у мышей (Suzuki et al., 2011). И наоборот, существует множество примеров, показывающих, что молекулярное вмешательство в механизмы стабилизации позвоночника, описанные выше, значительно влияет на когнитивные функции (Allen et al., 1998; Судхоф, 2008 г.; Редиес и др., 2012; Вукоевич и др., 2012; Ба и др., 2013).
Рисунок 2. Структурная пластичность астроцитов способствует поддержанию активированных синапсов . Левая панель: возбуждающие синапсы окружены астроцитарными отростками, демонстрирующими высокий уровень подвижности. Средняя панель: эта подвижность астроцитов регулируется высвобождаемым нейротрансмиттером, который активирует метаботропные рецепторы глутамата на астроцитарном отростке, что приводит к увеличению потока кальция и подвижности астроцитарного отростка.Правая панель: при управлении парадигмами, связанными с обучением (LTP, верхний синапс), эта усиленная подвижность приводит к усиленному и более стабильному покрытию синапса астроцитарным процессом, что приводит к длительной стабилизации синапса. Этот механизм может включать, среди прочего, вклад молекул адгезии.
Средняя панель: эта подвижность астроцитов регулируется высвобождаемым нейротрансмиттером, который активирует метаботропные рецепторы глутамата на астроцитарном отростке, что приводит к увеличению потока кальция и подвижности астроцитарного отростка.Правая панель: при управлении парадигмами, связанными с обучением (LTP, верхний синапс), эта усиленная подвижность приводит к усиленному и более стабильному покрытию синапса астроцитарным процессом, что приводит к длительной стабилизации синапса. Этот механизм может включать, среди прочего, вклад молекул адгезии.
Механизмы оборота позвоночника в зависимости от активности
Ставшие уже классическими эксперименты, проведенные Engert и Bonhoeffer (1999), предоставили первое доказательство того, что индукция пластичности может приводить к росту новых дендритных шипиков.Дальнейшие исследования нескольких групп подтвердили, что эти новые шипики демонстрируют синапсы со всеми характеристиками морфологически зрелых и функциональных контактов (Toni et al. , 1999; Nägerl et al., 2007). Исследования in vivo затем показали, что образование и удаление шипов происходит постоянно, в процессе развития и независимо от активности (Lendvai et al., 2000; Holtmaat et al., 2005; Zuo et al., 2005). Однако эти механизмы сильно усиливаются сенсорной или двигательной активностью (Holtmaat et al., 2006; Сюй и др., 2009). Постоянная тета-активность или индукция пластичности в срезах гиппокампа также сильно способствует обороту шипиков, усиливая как образование, так и устранение дендритных шипиков, что приводит к значимой зависимой от активности перенастройке цепи гиппокампа (De Roo et al., 2008b).
, 1999; Nägerl et al., 2007). Исследования in vivo затем показали, что образование и удаление шипов происходит постоянно, в процессе развития и независимо от активности (Lendvai et al., 2000; Holtmaat et al., 2005; Zuo et al., 2005). Однако эти механизмы сильно усиливаются сенсорной или двигательной активностью (Holtmaat et al., 2006; Сюй и др., 2009). Постоянная тета-активность или индукция пластичности в срезах гиппокампа также сильно способствует обороту шипиков, усиливая как образование, так и устранение дендритных шипиков, что приводит к значимой зависимой от активности перенастройке цепи гиппокампа (De Roo et al., 2008b).
Один интересный вопрос, поднятый этими наблюдениями, заключается в том, регулируются ли механизмы роста и ликвидации шипов независимо. Хотя активность, по-видимому, стимулирует оба процесса одновременно, некоторые недавние результаты свидетельствуют о том, что эти два механизма регулируются независимо друг от друга.Например, лечение эстрогенами приводит к значительному увеличению плотности позвоночника как в экспериментах in vitro , так и in vivo (Янкова и др.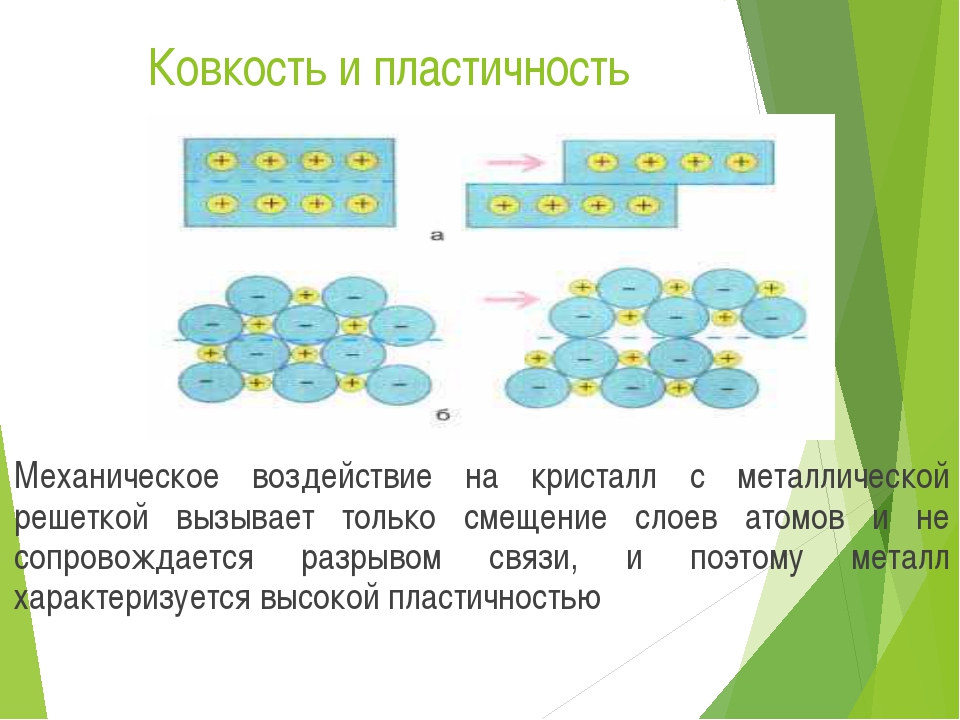 , 2001; Сакамото и др., 2003; Мендес и др., 2010b). Анализ оборота показал, что этот эффект возник в результате значительного увеличения механизмов роста шипов, но без изменений в удалении шипов, что объясняет изменения плотности. Точно так же специфические эффекты на механизмы роста шипов без изменений в удалении шипов наблюдались после интерференции с активированной киназой P21 (PAK3; Dubos et al., 2012), путем прямой активации цитоскелетного регулирующего белка сосудорасширяющего фосфопротеина (VASP; Lin et al., 2007; Nikonenko et al., 2013) и после применения BDNF или ингибитора фосфатазы и гомолога тензина (PTEN) BpV ( Танака и др., 2008; Бода и др., 2014). Следовательно, это указывает на то, что механизмы, которые способствуют полимеризации актина, могут непосредственно стимулировать рост позвоночника и что сигнальные пути, участвующие в регуляции синтеза белка, такие как путь PI3K-AKT-mTor, участвуют в контроле этого процесса роста.И наоборот, существуют также механизмы, которые избирательно воздействуют на элиминацию шипов, не нарушая процессов роста шипов.
, 2001; Сакамото и др., 2003; Мендес и др., 2010b). Анализ оборота показал, что этот эффект возник в результате значительного увеличения механизмов роста шипов, но без изменений в удалении шипов, что объясняет изменения плотности. Точно так же специфические эффекты на механизмы роста шипов без изменений в удалении шипов наблюдались после интерференции с активированной киназой P21 (PAK3; Dubos et al., 2012), путем прямой активации цитоскелетного регулирующего белка сосудорасширяющего фосфопротеина (VASP; Lin et al., 2007; Nikonenko et al., 2013) и после применения BDNF или ингибитора фосфатазы и гомолога тензина (PTEN) BpV ( Танака и др., 2008; Бода и др., 2014). Следовательно, это указывает на то, что механизмы, которые способствуют полимеризации актина, могут непосредственно стимулировать рост позвоночника и что сигнальные пути, участвующие в регуляции синтеза белка, такие как путь PI3K-AKT-mTor, участвуют в контроле этого процесса роста.И наоборот, существуют также механизмы, которые избирательно воздействуют на элиминацию шипов, не нарушая процессов роста шипов. Это может быть случай длительной депрессии, которая, как было показано в нескольких недавних исследованиях, приводит к уменьшению и исчезновению позвоночника (Becker et al., 2008; Oh et al., 2013; Wiegert and Oertner, 2013). Молекулярные механизмы, участвующие в этом эффекте, остаются неясными, но, вероятно, требуют активации метаботропных рецепторов как NMDA, так и глутамата (Oh et al., 2013). В соответствии с этой интерпретацией мы недавно определили особую роль включения субъединицы GluN3A в рецепторы NMDA в регуляции обрезки шипов (Kehoe et al., 2013). Субъединица GluN3A сильно экспрессируется в критические периоды развития, и ее экспрессия связана со снижением плотности шипиков (Roberts et al., 2009). Манипулируя экспрессией этой субъединицы с помощью экспериментов с усилением или потерей функции, мы обнаружили, что центральным механизмом, объясняющим эти эффекты, является избирательное увеличение обрезки шипов, связанное с экспрессией субъединицы GluN3A (Kehoe et al., 2014). Таким образом, эти результаты ясно указывают на то, что плотность позвоночника может регулироваться по-разному, вмешиваясь либо в механизмы роста позвоночника, либо в механизмы элиминации.
Это может быть случай длительной депрессии, которая, как было показано в нескольких недавних исследованиях, приводит к уменьшению и исчезновению позвоночника (Becker et al., 2008; Oh et al., 2013; Wiegert and Oertner, 2013). Молекулярные механизмы, участвующие в этом эффекте, остаются неясными, но, вероятно, требуют активации метаботропных рецепторов как NMDA, так и глутамата (Oh et al., 2013). В соответствии с этой интерпретацией мы недавно определили особую роль включения субъединицы GluN3A в рецепторы NMDA в регуляции обрезки шипов (Kehoe et al., 2013). Субъединица GluN3A сильно экспрессируется в критические периоды развития, и ее экспрессия связана со снижением плотности шипиков (Roberts et al., 2009). Манипулируя экспрессией этой субъединицы с помощью экспериментов с усилением или потерей функции, мы обнаружили, что центральным механизмом, объясняющим эти эффекты, является избирательное увеличение обрезки шипов, связанное с экспрессией субъединицы GluN3A (Kehoe et al., 2014). Таким образом, эти результаты ясно указывают на то, что плотность позвоночника может регулироваться по-разному, вмешиваясь либо в механизмы роста позвоночника, либо в механизмы элиминации.
До сих пор неясно, способствуют ли эти механизмы вращения позвоночника когнитивным функциям и каким образом. Однако сообщалось об изменениях оборота позвоночника в случаях умственной отсталости или расстройств аутистического спектра (Cruz-Martín et al., 2010; Pan et al., 2010; Dubos et al., 2012), и механизмы, которые регулируют рост и оборот позвоночника, также связаны с когнитивными дефектами и расстройствами аутистического спектра (Sharma et al., 2010; Cao et al., 2013; Boda et al., 2014).
Механизмы, лежащие в основе кластеризации позвоночника
Интересное наблюдение, относящееся к механизмам роста шипов, зависящим от активности, заключается в том, что образование новых шипов происходит неслучайным образом, но чаще наблюдается в непосредственной близости от активированных синапсов, что иногда приводит к образованию кластеров шипов (De Roo et al. др., 2008б; Дубос и др., 2012). Это явление, первоначально обнаруженное в культурах срезов гиппокампа, затем также наблюдалось в условиях in vivo после выполнения задания на обучение моторике (Fu et al. , 2012). Интригующий вопрос касается механизма, который может способствовать росту новых шипов в кластерах или в областях активности. Можно рассмотреть несколько гипотез. Во-первых, в работах in vitro есть данные о том, что применение глутамата или удаление глутамата из клетки на дендрите может быстро вызвать рост нового шипа (Richards et al., 2005; Квон и Сабатини, 2011). Показано, что это явление зависит от активности протеинкиназ постсинаптического дендрита. Таким образом, можно предположить, что интенсивная пресинаптическая активность и высвобождение глутамата могут вызвать рост новых шипиков в небольших областях мозга. Одной из основных трудностей с этой интерпретацией является время формирования позвоночника. Наблюдалось, что рост позвоночника происходит в течение нескольких минут после высвобождения глутамата, в то время как рост позвоночника, зависящий от активности, является гораздо более медленным процессом, который можно визуализировать только через несколько часов после стимуляции, спустя много времени после того, как произошло высвобождение глутамата (Engert and Bonhoeffer, 1999; De Roo et al.
, 2012). Интригующий вопрос касается механизма, который может способствовать росту новых шипов в кластерах или в областях активности. Можно рассмотреть несколько гипотез. Во-первых, в работах in vitro есть данные о том, что применение глутамата или удаление глутамата из клетки на дендрите может быстро вызвать рост нового шипа (Richards et al., 2005; Квон и Сабатини, 2011). Показано, что это явление зависит от активности протеинкиназ постсинаптического дендрита. Таким образом, можно предположить, что интенсивная пресинаптическая активность и высвобождение глутамата могут вызвать рост новых шипиков в небольших областях мозга. Одной из основных трудностей с этой интерпретацией является время формирования позвоночника. Наблюдалось, что рост позвоночника происходит в течение нескольких минут после высвобождения глутамата, в то время как рост позвоночника, зависящий от активности, является гораздо более медленным процессом, который можно визуализировать только через несколько часов после стимуляции, спустя много времени после того, как произошло высвобождение глутамата (Engert and Bonhoeffer, 1999; De Roo et al. ., 2008б). Хотя это не исключает роли высвобождения глутамата, это определенно указывает на то, что рост позвоночника, вызванный активностью, не эквивалентен эффектам высвобождения глутамата. Недавно была предложена другая интерпретация, основанная на экспериментах, анализирующих роль оксида азота (NO) в развитии синапсов (Никоненко и др., 2013). Оксид азота может образовываться в возбуждающих синапсах в ответ на активацию NMDA-рецептором NO-синтазы, которая тесно связана с постсинаптической плотностью (Burette et al., 2002; Исии и др., 2006; d’Anglemont de Tassigny et al., 2007). Вмешательства, которые блокировали продукцию NO или гуанилаткиназу, основного нижестоящего эффектора NO, сильно снижали механизмы роста позвоночника, включая усиление, вызванное индукцией пластичности (Никоненко и др., 2013). Наоборот, применение цГМФ усиливало образование шипов, подавляя эффекты синаптической активности. Интересно, что эти эффекты можно проследить до фосфорилирования регуляторного белка актина VASP киназой cGMP.
., 2008б). Хотя это не исключает роли высвобождения глутамата, это определенно указывает на то, что рост позвоночника, вызванный активностью, не эквивалентен эффектам высвобождения глутамата. Недавно была предложена другая интерпретация, основанная на экспериментах, анализирующих роль оксида азота (NO) в развитии синапсов (Никоненко и др., 2013). Оксид азота может образовываться в возбуждающих синапсах в ответ на активацию NMDA-рецептором NO-синтазы, которая тесно связана с постсинаптической плотностью (Burette et al., 2002; Исии и др., 2006; d’Anglemont de Tassigny et al., 2007). Вмешательства, которые блокировали продукцию NO или гуанилаткиназу, основного нижестоящего эффектора NO, сильно снижали механизмы роста позвоночника, включая усиление, вызванное индукцией пластичности (Никоненко и др., 2013). Наоборот, применение цГМФ усиливало образование шипов, подавляя эффекты синаптической активности. Интересно, что эти эффекты можно проследить до фосфорилирования регуляторного белка актина VASP киназой cGMP. Экспрессия фосфомутантов VASP, которые либо имитировали, либо предотвращали фосфорилирование киназой cGMP, воспроизводила или блокировала эффекты активности на механизмы роста шипов (Nikonenko et al., 2013). Более того, у мышей с дефицитом NO-синтазы развитие синапсов позвоночника было значительно нарушено, и воздействие на этих мышей обогащенной среды не способствовало увеличению плотности синапсов. Анализ распределения синапсов шипов также выявил отсутствие кластеризации синапсов, явление, легко наблюдаемое у мышей дикого типа (Никоненко и др., 2013). Таким образом, эти данные свидетельствуют о том, что один из механизмов, который может объяснить зависящее от активности скопление шипиков вокруг активных участков, может включать продукцию NO активированными синапсами, что, в свою очередь, стимулирует рост протрузий за счет фосфорилирования регуляторного белка VASP, способствуя таким образом образование синапсов в областях, где находятся активные партнеры (резюме на рисунке 3).
Экспрессия фосфомутантов VASP, которые либо имитировали, либо предотвращали фосфорилирование киназой cGMP, воспроизводила или блокировала эффекты активности на механизмы роста шипов (Nikonenko et al., 2013). Более того, у мышей с дефицитом NO-синтазы развитие синапсов позвоночника было значительно нарушено, и воздействие на этих мышей обогащенной среды не способствовало увеличению плотности синапсов. Анализ распределения синапсов шипов также выявил отсутствие кластеризации синапсов, явление, легко наблюдаемое у мышей дикого типа (Никоненко и др., 2013). Таким образом, эти данные свидетельствуют о том, что один из механизмов, который может объяснить зависящее от активности скопление шипиков вокруг активных участков, может включать продукцию NO активированными синапсами, что, в свою очередь, стимулирует рост протрузий за счет фосфорилирования регуляторного белка VASP, способствуя таким образом образование синапсов в областях, где находятся активные партнеры (резюме на рисунке 3).
Рисунок 3. Регуляция локального роста позвоночника с помощью оксида азота (NO) .Левая панель: нейрональная NO-синтаза (nNOS), присутствующая в постсинаптической плотности и тесно связанная с рецепторами PSD-95 и NMDA, генерирует NO при активации рецептора NMDA и притоке кальция. Средняя панель: NO, генерируемый LTP в активированных синапсах, локально диффундирует и запускает активацию цГМФ-зависимого каскада, приводящего к фосфорилированию регуляторного белка цитоскелета VASP с помощью цГМФ-зависимой киназы. Это, в свою очередь, способствует удлинению актиновых филаментов и образованию шипиков. Правая панель: этот механизм позволяет формировать кластеры шипиковых синапсов вокруг активированных контактов, явление, отсутствующее у мышей с дефицитом nNOS.Электронно-микроскопическое изображение иллюстрирует скопления дендритных шипов (закрашенных розой) в гиппокампе мыши дикого типа, выращенной в обогащенной среде.
Регуляция локального роста позвоночника с помощью оксида азота (NO) .Левая панель: нейрональная NO-синтаза (nNOS), присутствующая в постсинаптической плотности и тесно связанная с рецепторами PSD-95 и NMDA, генерирует NO при активации рецептора NMDA и притоке кальция. Средняя панель: NO, генерируемый LTP в активированных синапсах, локально диффундирует и запускает активацию цГМФ-зависимого каскада, приводящего к фосфорилированию регуляторного белка цитоскелета VASP с помощью цГМФ-зависимой киназы. Это, в свою очередь, способствует удлинению актиновых филаментов и образованию шипиков. Правая панель: этот механизм позволяет формировать кластеры шипиковых синапсов вокруг активированных контактов, явление, отсутствующее у мышей с дефицитом nNOS.Электронно-микроскопическое изображение иллюстрирует скопления дендритных шипов (закрашенных розой) в гиппокампе мыши дикого типа, выращенной в обогащенной среде.
Хотя функциональное значение этого феномена кластеризации до сих пор неясно, интересно, что вмешательство в продукцию NO или передачу сигналов связано с нарушением когнитивных функций и социальной дисфункцией (Kirchner et al. , 2004; Kelley et al., 2009; Tanda et al. ., 2009; Wass et al., 2009) и варианты гена NO-синтазы 1 связаны с шизофренией (Shinkai et al., 2002; Бернштейн и др., 2011).
, 2004; Kelley et al., 2009; Tanda et al. ., 2009; Wass et al., 2009) и варианты гена NO-синтазы 1 связаны с шизофренией (Shinkai et al., 2002; Бернштейн и др., 2011).
Синаптические дефекты при нарушениях психики, связанных с развитием
Все эти данные подчеркивают сложность механизмов, регулирующих динамику возбуждающих синапсов во время развития, и убедительно свидетельствуют о том, что любое вмешательство в эти свойства структурной пластичности может иметь значительное влияние на организацию и функциональные свойства возбуждающих сетей. Можно ожидать, что изменения в стабильности позвоночника или обороте позвоночника повлияют на количество, а также на специфичность возбуждающих синапсов, что приведет к формированию синаптических цепей с различными аномальными характеристиками, такими как гипер- или гипосвязность и гипер- или гиподинамические реакции на активность (Mukai et al., 2008; Пан и др., 2010; Цю и др., 2011). Все эти различные изменения, влияя на развитие и функциональные свойства синаптических сетей, могут снижать надежность обработки информации и приводить к формированию дезадаптирующих мозговых цепей (Sigurdsson et al. , 2010). В зависимости от вовлеченного молекулярного механизма изменения могут по-разному влиять на различные области мозга или оказывать большее влияние на разных фазах развития, тем самым объясняя вариабельность клинических или поведенческих фенотипов.Можно было бы также ожидать, что изменения структурной пластичности с сильным воздействием в критические периоды развития могут иметь более разрушительные последствия, которые трудно обратить вспять и которые потребуют очень раннего вмешательства для компенсации дефектов. Такие ранние подходы в настоящее время тестируются во многих ситуациях, связанных с расстройствами аутистического спектра (Canitano, 2014).
, 2010). В зависимости от вовлеченного молекулярного механизма изменения могут по-разному влиять на различные области мозга или оказывать большее влияние на разных фазах развития, тем самым объясняя вариабельность клинических или поведенческих фенотипов.Можно было бы также ожидать, что изменения структурной пластичности с сильным воздействием в критические периоды развития могут иметь более разрушительные последствия, которые трудно обратить вспять и которые потребуют очень раннего вмешательства для компенсации дефектов. Такие ранние подходы в настоящее время тестируются во многих ситуациях, связанных с расстройствами аутистического спектра (Canitano, 2014).
Основываясь на этих наблюдениях, мы предполагаем, что возможной общей чертой различных психических расстройств, связанных с развитием, может быть наличие изменений в динамических свойствах возбуждающих синапсов.Это согласуется с сильным компонентом развития этих нарушений и множеством возможных синаптических белков. Это также может объяснить тот факт, что области, показывающие высокую степень пластичности, такие как префронтальная кора и гиппокамп, очень часто демонстрируют функциональные дефекты. Наконец, в соответствии с этой гипотезой, сигнальные пути, играющие важную роль в регуляции динамики позвоночника, являются основными кандидатами на участие в психических расстройствах.Примеры включают передачу сигналов Rho GTPases и их регуляцию актинового цитоскелета и сигнальных каскадов, участвующих в контроле синтеза белка (Sawicka and Zukin, 2012; Ba et al., 2013). Эти две сигнальные системы особенно важны в возбуждающих синапсах для контроля морфологии и пластичности позвоночника, а некоторые генетические изменения, связанные с психическими расстройствами, влияют на молекулы, участвующие в этих двух путях (Boda et al., 2010; Fromer et al., 2014).
Наконец, в соответствии с этой гипотезой, сигнальные пути, играющие важную роль в регуляции динамики позвоночника, являются основными кандидатами на участие в психических расстройствах.Примеры включают передачу сигналов Rho GTPases и их регуляцию актинового цитоскелета и сигнальных каскадов, участвующих в контроле синтеза белка (Sawicka and Zukin, 2012; Ba et al., 2013). Эти две сигнальные системы особенно важны в возбуждающих синапсах для контроля морфологии и пластичности позвоночника, а некоторые генетические изменения, связанные с психическими расстройствами, влияют на молекулы, участвующие в этих двух путях (Boda et al., 2010; Fromer et al., 2014).
Сигнальный путь Rho GTPases включает несколько молекул, которые связаны с умственной отсталостью, расстройствами аутистического спектра или шизофренией.Примеры включают мембранные рецепторы и молекулы адгезии, способные активировать Rho-GTPases (кадгерины, эфрины), регуляторы Rho-GTPases (oligophrenin1, GDI1, ARHGEF6, EPAC2, CNK2 (коннектор-энхансер KSR-2), srGAP3, Disc1, SynGAP), эффекторы Rho GTPases (PAK3), а также регуляторные белки цитоскелета (Endris et al. , 2002; Govek et al., 2004; Nodé-Langlois et al., 2006; Woolfrey et al., 2009; Hayashi-Takagi et al., 2010; Клемент и др., 2012; Лим и др., 2014). Во многих из этих случаев сообщалось об изменениях морфологии или функции синапсов, хотя анализ динамики шипиков все еще часто отсутствует.Есть один пример, который мы изучили более подробно и который касается белка умственной отсталости PAK3. Несколько мутаций PAK3 связаны с умственной отсталостью у людей. Было обнаружено, что экспрессия мутантного PAK3 в нейронах гиппокампа или генетическая и фармакологическая супрессия PAK3 приводят к изменениям морфологии и динамики шипов (Kreis et al., 2007; Dubos et al., 2012). Двумя основными особенностями, наблюдаемыми после вмешательства в функцию PAK3, были усиление механизмов спонтанного роста позвоночника и блок стабилизации позвоночника, зависящей от активности.Следовательно, это приводит к чрезмерно динамичной синаптической сети, в которой активность не способна избирательно стабилизировать активированные синапсы (Dubos et al.
, 2002; Govek et al., 2004; Nodé-Langlois et al., 2006; Woolfrey et al., 2009; Hayashi-Takagi et al., 2010; Клемент и др., 2012; Лим и др., 2014). Во многих из этих случаев сообщалось об изменениях морфологии или функции синапсов, хотя анализ динамики шипиков все еще часто отсутствует.Есть один пример, который мы изучили более подробно и который касается белка умственной отсталости PAK3. Несколько мутаций PAK3 связаны с умственной отсталостью у людей. Было обнаружено, что экспрессия мутантного PAK3 в нейронах гиппокампа или генетическая и фармакологическая супрессия PAK3 приводят к изменениям морфологии и динамики шипов (Kreis et al., 2007; Dubos et al., 2012). Двумя основными особенностями, наблюдаемыми после вмешательства в функцию PAK3, были усиление механизмов спонтанного роста позвоночника и блок стабилизации позвоночника, зависящей от активности.Следовательно, это приводит к чрезмерно динамичной синаптической сети, в которой активность не способна избирательно стабилизировать активированные синапсы (Dubos et al. , 2012). Это также отражалось в присутствии высокой доли тонких незрелых шипов и сниженном количестве крупных шипов грибовидного типа (Boda et al., 2004). Таким образом, можно предположить, что эти дефекты привели к неспособности поддерживать важные соединения и развивать специфику в организации проводки.
, 2012). Это также отражалось в присутствии высокой доли тонких незрелых шипов и сниженном количестве крупных шипов грибовидного типа (Boda et al., 2004). Таким образом, можно предположить, что эти дефекты привели к неспособности поддерживать важные соединения и развивать специфику в организации проводки.
Второй важный путь, тесно связанный с психическими расстройствами, включает ряд молекул, регулирующих локальный синтез белка.Сюда входят мембранные рецепторы (BDNF и TrkB, рецептор инсулина, эфрины) или молекулы адгезии и каркаса (интегрины, комплекс нейролигин/нейрексин, хвостовик, SynGAP), внутриклеточные медиаторы, участвующие в передаче сигналов mTOR (PI3K, Akt, PTEN, TSC2), и регуляторы синтез белка (FMRP, CYFIP1) (Kumar et al., 2005; Lai and Ip, 2009; Mendez et al., 2010a; Sharma et al., 2010; Cuesto et al., 2011; Clement et al., 2012; Boda и др., 2014). Этот путь также играет важную роль в регуляции силы синапсов, и в большинстве случаев сообщалось об изменениях морфологии или плотности шипиков, что предполагает нарушение структурной пластичности. Одним из наиболее изученных примеров, безусловно, является синдром Fragile X, связанный с множественными повторами CGG в гене fmr1 (Bhakar et al., 2012). У мышей с дефицитом белка FMRP обнаруживаются изменения морфологии шипов с увеличением доли тонких удлиненных шипов (Comery et al., 1997, но см. также Wijetunge et al., 2014). Недавняя работа с использованием мышей с нокаутом Fmr1 показала, что эти шипы гипердинамичны, не реагируют на сенсорную стимуляцию (Pan et al., 2010) и не обладают стабильностью (Cruz-Martín et al., 2010). В экспериментах с культивированием срезов мы также обнаружили, что оборот шипов был чрезмерно чувствителен к активности и, более конкретно, индукция пластичности не вызывала дифференциальной стабилизации активированных шипов. Интересно, что в этих экспериментах нам удалось обратить этот дефицит за счет применения ингибитора PTEN, который усиливает сигнальный путь PI3K-Akt-mTOR (Boda et al., 2014), в то время как вмешательство в метаботропные рецепторы глутамата было лишь частично эффективным.
Одним из наиболее изученных примеров, безусловно, является синдром Fragile X, связанный с множественными повторами CGG в гене fmr1 (Bhakar et al., 2012). У мышей с дефицитом белка FMRP обнаруживаются изменения морфологии шипов с увеличением доли тонких удлиненных шипов (Comery et al., 1997, но см. также Wijetunge et al., 2014). Недавняя работа с использованием мышей с нокаутом Fmr1 показала, что эти шипы гипердинамичны, не реагируют на сенсорную стимуляцию (Pan et al., 2010) и не обладают стабильностью (Cruz-Martín et al., 2010). В экспериментах с культивированием срезов мы также обнаружили, что оборот шипов был чрезмерно чувствителен к активности и, более конкретно, индукция пластичности не вызывала дифференциальной стабилизации активированных шипов. Интересно, что в этих экспериментах нам удалось обратить этот дефицит за счет применения ингибитора PTEN, который усиливает сигнальный путь PI3K-Akt-mTOR (Boda et al., 2014), в то время как вмешательство в метаботропные рецепторы глутамата было лишь частично эффективным. Круз-Мартин и др., 2010; Бода и др., 2014). Этот ингибитор PTEN также улучшал связанный с этим дефицит механизмов LTP, а при введении мышам с дефицитом FMR1 восстанавливал обратное обучение в задаче водного лабиринта Морриса (Boda et al., 2014). Хотя многое еще предстоит понять в отношении механизмов, с помощью которых локальный синтез белка может влиять на структурную пластичность, эти данные предполагают, что эта система может быть интересной мишенью для модулирования свойств динамики позвоночника и, в конечном итоге, уравновешивать некоторые дефициты, наблюдаемые при ассоциированных расстройствах.
Круз-Мартин и др., 2010; Бода и др., 2014). Этот ингибитор PTEN также улучшал связанный с этим дефицит механизмов LTP, а при введении мышам с дефицитом FMR1 восстанавливал обратное обучение в задаче водного лабиринта Морриса (Boda et al., 2014). Хотя многое еще предстоит понять в отношении механизмов, с помощью которых локальный синтез белка может влиять на структурную пластичность, эти данные предполагают, что эта система может быть интересной мишенью для модулирования свойств динамики позвоночника и, в конечном итоге, уравновешивать некоторые дефициты, наблюдаемые при ассоциированных расстройствах.
Заключение
Функциональные свойства синаптической пластичности за счет быстрого изменения синаптической силы позволяют быстро адаптировать сетевую активность, которая имеет решающее значение для обработки информации. Однако в долгосрочной перспективе свойства структурной пластичности могут обеспечить более значительную и стабильную перестройку синаптических сетей как за счет образования новых связей, так и за счет стабилизации специфических контактов. Эти свойства структурной пластичности особенно важны во время развития, когда они способствуют формированию структурной организации мозговых цепей посредством деятельности.Молекулярный анализ этих структурных свойств начал идентифицировать ключевые сигнальные пути, участвующие в этих синаптических реорганизациях, которые также, по-видимому, являются сильными кандидатами на участие в когнитивных и психических расстройствах. Следовательно, общий знаменатель нарушений развития может включать изменения в динамике позвоночника, которые влияют на связность и специфичность мозговых цепей. Более систематический анализ этих свойств и их функциональных последствий должен позволить лучше понять, как они влияют на обработку информации, и это может в конечном итоге привести к новым возможностям лечения этих расстройств.
Эти свойства структурной пластичности особенно важны во время развития, когда они способствуют формированию структурной организации мозговых цепей посредством деятельности.Молекулярный анализ этих структурных свойств начал идентифицировать ключевые сигнальные пути, участвующие в этих синаптических реорганизациях, которые также, по-видимому, являются сильными кандидатами на участие в когнитивных и психических расстройствах. Следовательно, общий знаменатель нарушений развития может включать изменения в динамике позвоночника, которые влияют на связность и специфичность мозговых цепей. Более систематический анализ этих свойств и их функциональных последствий должен позволить лучше понять, как они влияют на обработку информации, и это может в конечном итоге привести к новым возможностям лечения этих расстройств.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Работа выполнена при поддержке Швейцарского научного фонда (грант 310030B_144080) и NCCR Synapsy.
Ссылки
Аллен, К. М., Глисон, Дж. Г., Багродиа, С., Партингтон, М. В., Макмиллан, Дж. К., Серионе, Р.А. и др. (1998). Мутация PAK3 при несиндромальной Х-сцепленной умственной отсталости. Нац. Жене. 20, 25–30. дои: 10.1038/1675
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Беккер Н., Виренга С. Дж., Фонсека Р., Бонхёффер Т. и Нэгерл У. В. (2008). Индукция LTD вызывает морфологические изменения пресинаптических бутонов и уменьшает их контакты с шипиками. Нейрон 60, 590–597. doi: 10.1016/j.neuron.2008.09.018
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Бернардинелли, Ю., Randall, J., Janett, E., Nikonenko, I., Konig, S., Jones, E.V., et al. (2014). Зависимая от активности структурная пластичность перисинаптических астроцитарных доменов способствует стабильности возбуждающих синапсов. Курс. биол. 24, 1679-1688. doi: 10.1016/j.cub.2014.06.025
Курс. биол. 24, 1679-1688. doi: 10.1016/j.cub.2014.06.025
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Бернштейн, Х.Г., Кейлхофф, Г., Штайнер, Дж., Добровольный, Х., и Богертс, Б. (2011). Окись азота и шизофрения: современные знания и новые концепции терапии. Нейрол ЦНС. Беспорядок. Наркотики 10, 792–807. дои: 10.2174/187152711798072392
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Бода Б., Альбери С., Никоненко И., Нод-Ланглуа Р., Журден П., Моосмайер М. и соавт. (2004). Белок умственной отсталости PAK3 способствует образованию синапсов и пластичности в гиппокампе. J. Neurosci. 24, 10816–10825. doi: 10.1523/jneurosci.2931-04.2004
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Бода, Б., Мендес П., Бури-Джамот Б., Магара Ф. и Мюллер Д. (2014). Изменение динамики позвоночника, опосредованной активностью, и ухудшение способности к обучению в мышиной модели синдрома Fragile X. евро. Дж. Нейроски. 39, 11:30–11:37. doi: 10.1111/ejn.12488
(2014). Изменение динамики позвоночника, опосредованной активностью, и ухудшение способности к обучению в мышиной модели синдрома Fragile X. евро. Дж. Нейроски. 39, 11:30–11:37. doi: 10.1111/ejn.12488
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Bozdagi, O., Wang, X.B., Nikitczuk, J.S., Anderson, T.R., Bloss, E.B., Radice, G.L., et al. (2010). Для сохранения скоординированной долговременной потенциации и увеличения дендритных шипиков в зрелых синапсах CA1 гиппокампа требуется N-кадгерин. J. Neurosci. 30, 9984–9989. doi: 10.1523/jneurosci.1223-10.2010
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Бюрет, А., Забель, У., Вайнберг, Р. Дж., Шмидт, Х. Х., и Вальчанофф, Дж. Г. (2002). Синаптическая локализация синтазы оксида азота и растворимой гуанилатциклазы в гиппокампе. J. Neurosci. 22, 8961–8970.
Опубликовано Резюме | Опубликован полный текст | Академия Google
Цао, К. , Rioult-Pedotti, M.S., Migani, P., Yu, C.J., Tiwari, R., Parang, K., et al. (2013). Нарушение передачи сигналов TrkB-PSD-95 при синдроме Ангельмана. PLoS Биол. 11:e1001478. doi: 10.1371/journal.pbio.1001848
, Rioult-Pedotti, M.S., Migani, P., Yu, C.J., Tiwari, R., Parang, K., et al. (2013). Нарушение передачи сигналов TrkB-PSD-95 при синдроме Ангельмана. PLoS Биол. 11:e1001478. doi: 10.1371/journal.pbio.1001848
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Чен, Л.Ю., Рекс, К.С., Казале, М.С., Галл, К.М., и Линч, Г. (2007). Изменения синаптической морфологии сопровождают передачу сигналов актином во время LTP. J. Neurosci. 27, 5363–5372.doi: 10.1523/jneurosci.0164-07.2007
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Клемент, Дж. П., Ацети, М., Кресон, Т. К., Озкан, Э. Д., Ши, Ю., Рейш, Н. Дж., и соавт. (2012). Патогенные мутации SYNGAP1 нарушают когнитивное развитие, нарушая созревание синапсов дендритных шипов. Сотовый 151, 709–723. doi: 10.1016/j.cell.2012.08.045
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Коми, Т. A., Harris, J.B., Willems, P.J., Oostra, B.A., Irwin, S.A., Weiler, I.J., et al. (1997). Аномальные дендритные шипы у хрупких мышей с нокаутом X: дефицит созревания и обрезки. Проц. Натл. акад. науч. США 94, 5401–5404. doi: 10.1073/pnas.94.10.5401
A., Harris, J.B., Willems, P.J., Oostra, B.A., Irwin, S.A., Weiler, I.J., et al. (1997). Аномальные дендритные шипы у хрупких мышей с нокаутом X: дефицит созревания и обрезки. Проц. Натл. акад. науч. США 94, 5401–5404. doi: 10.1073/pnas.94.10.5401
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Куесто Г., Энрикес-Баррето Л., Карамес К., Кантареро М., Гасулл Х., Санди К. и др. (2011). Активация фосфоинозитид-3-киназы контролирует синаптогенез и спиногенез в нейронах гиппокампа. J. Neurosci. 31, 2721–2733. doi: 10.1523/JNEUROSCI.4477-10.2011
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
d’Anglemont de Tassigny, X., Campagne, C., Dehouck, B., Leroy, D., Holstein, G.R., Beauvillain, J.C., et al. (2007). Связывание нейрональной синтазы оксида азота с рецепторами NMDA через постсинаптическую плотность-95 зависит от эстрогена и способствует центральному контролю репродукции взрослых самок. J. Neurosci. 27, 6103–6114.doi: 10.1523/jneurosci.5595-06.2007
J. Neurosci. 27, 6103–6114.doi: 10.1523/jneurosci.5595-06.2007
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Де Ру М., Клаузер П., Мендес П., Полья Л. и Мюллер Д. (2008a). Зависимое от активности формирование PSD и стабилизация вновь образованных шипиков в культурах срезов гиппокампа. Церебр. Кортекс 18, 151–161. doi: 10.1093/cercor/bhm041
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Дюбо, А., Комбо, Г., Bernardinelli Ю., Барнье, Дж В., Хартли, О., Гэртнер, H., и др. (2012). Изменение динамики синаптической сети белком умственной отсталости PAK3. J. Neurosci. 32, 519–527. DOI: 10.1523 / JNEUROSCI.3252-11.2012
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Эндрис В., Вогатцки Б., Леймер У., Барч Д., Затыка М., Латиф Ф. и соавт. (2002). Новый Rho-ГТФ активации ген MEGAP / srGAP3 имеет предполагаемую роль в тяжелой умственной отсталости. Проц. Натл. акад. науч. U S A 99, 11754-11759. DOI: 10.1073 / pnas.162241099
Новый Rho-ГТФ активации ген MEGAP / srGAP3 имеет предполагаемую роль в тяжелой умственной отсталости. Проц. Натл. акад. науч. U S A 99, 11754-11759. DOI: 10.1073 / pnas.162241099
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Фромер, М., Поклингтон, А. И., Кавана, Д. Х. Уильямс, Х. Дж, Dwyer, С., Гормли, P. и др. (2014). De Novo мутации в шизофрению подразумевающих синаптических сетей. Природа 506, 179-184. DOI: 10.1038 / nature12929
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Fukazawa, Ю.В., Сайтох Ю., Одзава Ф., Охта Ю., Мидзуно К. и Инокути К. (2003). ДП гиппокампа сопровождается повышенным содержанием F-актина в дендритных шипах, что необходимо для поддержания позднего ДП in vivo. Нейрон 38, 447–460. doi: 10.1016/s0896-6273(03)00206-x
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Говек, Э. Э., Ньюи, С. Э., Акерман, С. Дж., Кросс, Дж. Р., Ван дер Векен, Л., и Ван Элст, Л. (2004). Х-сцепленный белок умственной отсталости олигофренин-1 необходим для морфогенеза дендритных шипов. Нац. Неврологи. 7, 364–372. doi: 10.1038/nn1210
Э., Ньюи, С. Э., Акерман, С. Дж., Кросс, Дж. Р., Ван дер Векен, Л., и Ван Элст, Л. (2004). Х-сцепленный белок умственной отсталости олигофренин-1 необходим для морфогенеза дендритных шипов. Нац. Неврологи. 7, 364–372. doi: 10.1038/nn1210
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Хаяси-Такаги, А., Такаки, М., Грациане, Н., Сешадри, С., Мердок, Х., Данлоп, А. Дж., и соавт. (2010). Disrupted-in-Schizophrenia 1 (DISC1) регулирует шипы глутаматного синапса через Rac1. Нац. Неврологи. 13, 327–332. doi: 10.1038/nn.2487
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Холтмаат, А.J., Trachtenberg, J.T., Wilbrecht, L., Shepherd, G.M., Zhang, X., Knott, G.W., et al. (2005). Временные и постоянные дендритные шипы в неокортексе in vivo. Нейрон 45, 279–291. doi: 10.1016/j.neuron.2005.01.003
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Холтмаат А. , Уилбрехт Л., Нотт Г.В., Велкер Э. и Свобода К. (2006). Зависимый от опыта и специфичный для типа клеток рост шипов в неокортексе. Природа 441, 979–983.doi: 10.1038/nature04783
, Уилбрехт Л., Нотт Г.В., Велкер Э. и Свобода К. (2006). Зависимый от опыта и специфичный для типа клеток рост шипов в неокортексе. Природа 441, 979–983.doi: 10.1038/nature04783
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Хонкура, Н., Мацудзаки, М., Ногучи, Дж., Эллис-Дэвис, Г.К., и Касаи, Х. (2008). Подшиповая организация актиновых волокон регулирует структуру и пластичность дендритных шипиков. Нейрон 57, 719–729. doi: 10.1016/j.neuron.2008.01.013
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Исии, Х., Сибуя, К., Ohta, Y., Mukai, H., Uchino, S., Takata, N., et al. (2006). Усиление продукции оксида азота за счет ассоциации синтазы оксида азота с рецепторами N-метил-D-аспартата посредством постсинаптической плотности 95 в генетически модифицированных клетках яичника китайского хомячка: визуализация флуоресценции в реальном времени с использованием чувствительного к оксиду азота красителя.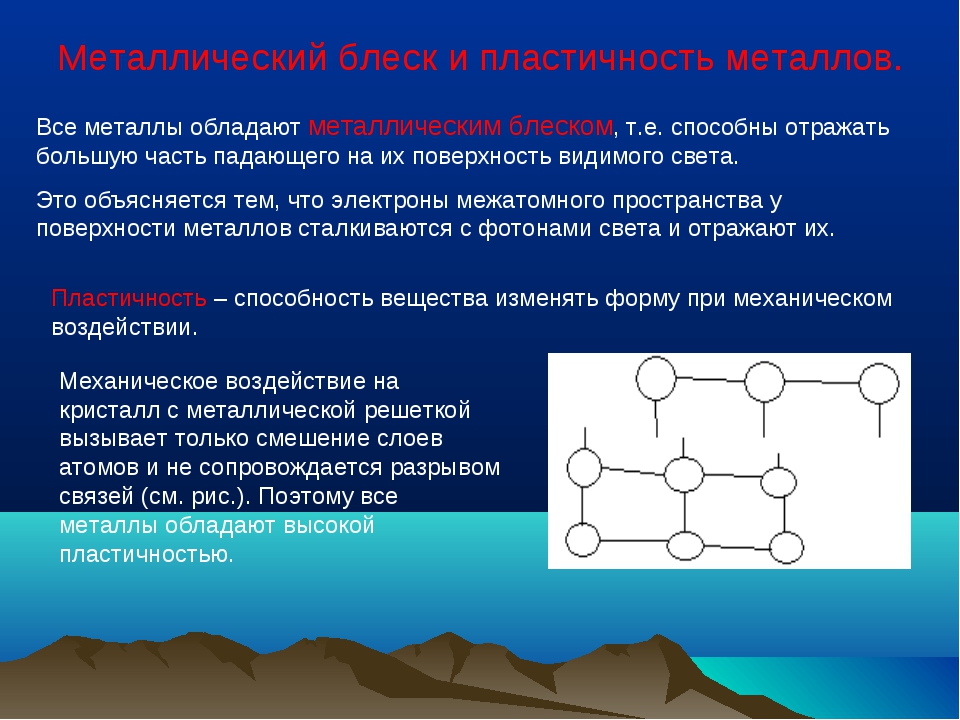 Дж. Нейрохим. 96, 1531–1539. doi: 10.1111/j.1471-4159.2006.03656.x
Дж. Нейрохим. 96, 1531–1539. doi: 10.1111/j.1471-4159.2006.03656.x
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Касаи, Х., Мацудзаки М., Ногучи Дж., Ясумацу Н. и Накахара Х. (2003). Отношения структура-стабильность-функция дендритных шипиков. Trends Neurosci. 26, 360–368. doi: 10.1016/s0166-2236(03)00162-0
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Кек Т., Мрсик-Флогель Т. Д., Ваз Афонсо М., Эйзель У. Т., Бонхёффер Т. и Хюбенер М. (2008). Массивная реструктуризация нейронных цепей во время функциональной реорганизации зрительной коры взрослого человека. Нац. Неврологи. 11, 1162–1167. doi: 10.1038/nn.2181
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Кехо, Л. А., Беллоне, К., Де Ру, М., Зандуэта, А., Дей, П. Н., Перес-Отаньо, И., и др. (2014). GluN3A способствует сокращению и дестабилизации дендритных шипов во время постнатального развития. J. Neurosci. 34, 9213–9221. doi: 10.1523/JNEUROSCI.5183-13.2014
(2014). GluN3A способствует сокращению и дестабилизации дендритных шипов во время постнатального развития. J. Neurosci. 34, 9213–9221. doi: 10.1523/JNEUROSCI.5183-13.2014
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Киршнер, Л., Weitzdoerfer, R., Hoeger, H., Url, A., Schmidt, P., Engelmann, M., et al. (2004). Нарушение когнитивных функций у мышей с нокаутом нейрональной синтазы оксида азота связано с нарушениями белков гиппокампа. Оксид азота 11, 316–330. doi: 10.1016/j.niox.2004.10.005
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Крайс П., Тевено Э., Руссо В., Бода Б., Мюллер Д. и Барнье Дж. В. (2007). Активируемая р21 киназа 3, участвующая в умственной отсталости, регулирует морфогенез позвоночника посредством Cdc42-зависимого пути. Дж. Биол. хим. 282, 21497–21506. дои: 10.1074/jbc.m703298200
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Кумар, В. , Чжан, М. Х., Суонк, М. В., Кунц, Дж., и Ву, Г. Ю. (2005). Регуляция морфогенеза дендритов сигнальными путями Ras-PI3K-Akt-mTOR и Ras-MAPK. J. Neurosci. 25, 11288–11299. doi: 10.1523/jneurosci.2284-05.2005
, Чжан, М. Х., Суонк, М. В., Кунц, Дж., и Ву, Г. Ю. (2005). Регуляция морфогенеза дендритов сигнальными путями Ras-PI3K-Akt-mTOR и Ras-MAPK. J. Neurosci. 25, 11288–11299. doi: 10.1523/jneurosci.2284-05.2005
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Мендес, П., Де Ру М., Полья Л., Клаузер П. и Мюллер Д. (2010a). N-кадгерин обеспечивает долговременную стабилизацию позвоночника, вызванную пластичностью. J. Cell Biol. 189, 589–600. doi: 10.1083/jcb.201003007
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Mukai, J., Dhilla, A., Drew, L.J., Stark, K.L., Cao, L., MacDermott, A.B., et al. (2008). Зависимый от пальмитоилирования дефицит развития нервной системы в мышиной модели микроделеции 22q11. Нац.Неврологи. 11, 1302–1310. doi: 10.1038/nn.2204
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Нэгерл, У.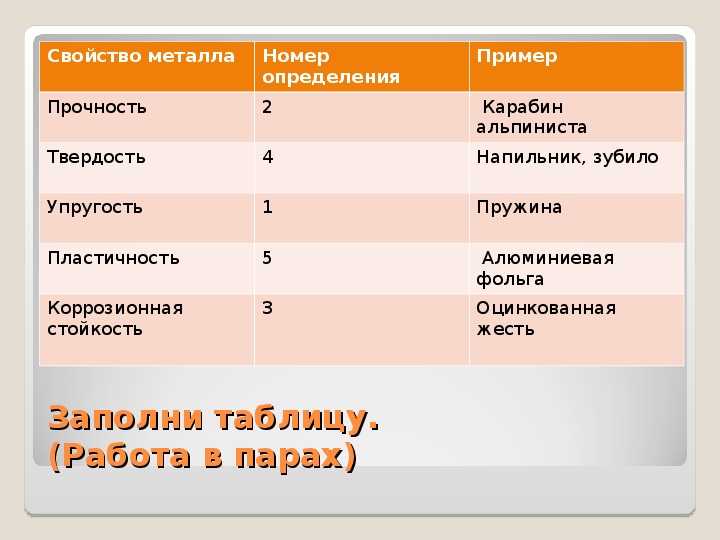 В., Кёстингер, Г., Андерсон, Дж. К., Мартин, К. А., и Бонхёффер, Т. (2007). Затяжной синаптогенез после зависимого от активности спиногенеза в нейронах гиппокампа. J. Neurosci. 27, 8149–8156. doi: 10.1523/JNEUROSCI.0511-07.2007
В., Кёстингер, Г., Андерсон, Дж. К., Мартин, К. А., и Бонхёффер, Т. (2007). Затяжной синаптогенез после зависимого от активности спиногенеза в нейронах гиппокампа. J. Neurosci. 27, 8149–8156. doi: 10.1523/JNEUROSCI.0511-07.2007
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Никоненко И., Никоненко А., Мендес П., Мичурина Т. В., Ениколопов Г. и Мюллер Д. (2013). Оксид азота опосредует локальное развитие возбуждающих синапсов, зависящее от активности. Проц. Натл. акад. науч. США 110, E4142–E4151. doi: 10.1073/pnas.1311927110
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Пан Ф., Олдридж Г. М., Гриноу В. Т. и Ган В. Б. (2010). Дендритная нестабильность позвоночника и нечувствительность к модуляции сенсорным опытом в мышиной модели синдрома ломкой Х-хромосомы. Проц. Натл. акад. науч. США 107, 17768–17773. doi: 10.1073/pnas.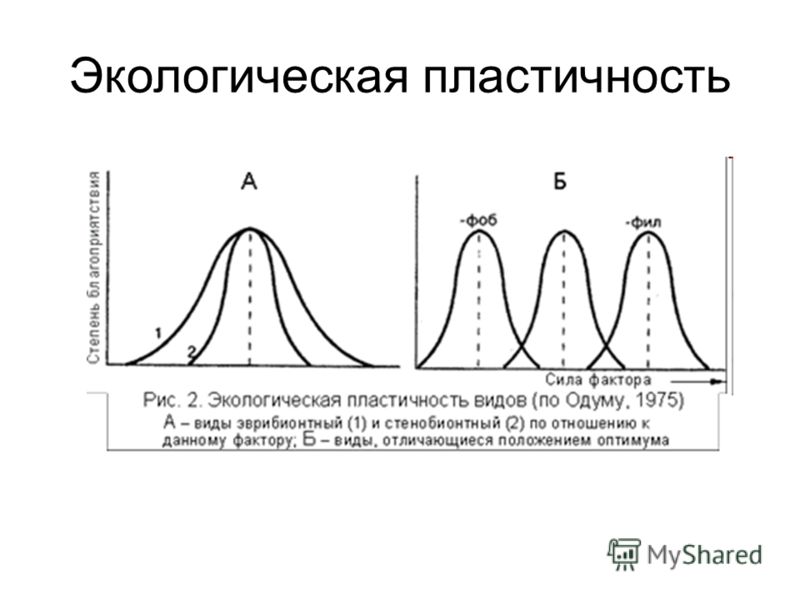 1012496107
1012496107
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Цю, С., Андерсон, К.Т., Левитт, П., и Шеперд, Г.М. (2011). Схемно-специфическая внутрикортикальная гиперсвязность у мышей с делецией тирозинкиназы рецептора Met, связанной с аутизмом. J. Neurosci. 31, 5855–5864. doi: 10.1523/JNEUROSCI.6569-10.2011
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Рекс, К.С., Чен, Л.Ю., Шарма, А., Лю, Дж., Бабаян, А.Х., Галл, С.М., и соавт. (2009). Различные сигнальные пути, зависимые от Rho GTPase, инициируют последовательные шаги в консолидации долговременного потенцирования. J. Cell Biol. 186, 85–97. doi: 10.1083/jcb.200
4
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Ричардс, Д. А., Матеос, Дж. М., Хьюгель, С., де Паола, В., Карони, П., Гавилер, Б. Х., и соавт. (2005). Глутамат индуцирует быстрое образование выступов головки позвоночника в культурах срезов гиппокампа. Проц. Натл. акад. науч. США 102, 6166–6171. doi: 10.1073/pnas.0501881102
(2005). Глутамат индуцирует быстрое образование выступов головки позвоночника в культурах срезов гиппокампа. Проц. Натл. акад. науч. США 102, 6166–6171. doi: 10.1073/pnas.0501881102
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Робертс А.С., Диес-Гарсия Дж., Родригис Р.М., Лопес И.П., Лухан Р., Мартинес-Турильяс Р. и др. (2009). Понижающая регуляция NR3A-содержащих NMDAR необходима для созревания синапсов и консолидации памяти. Нейрон 63, 342–356. doi: 10.1016/j.neuron.2009.06.016
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Сакамото, Х., Мезаки Ю., Шикими Х., Укена К. и Цуцуи К. (2003). Рост дендритов и образование шипов в ответ на эстроген в развивающихся клетках Пуркинье. Эндокринология 144, 4466–4477. doi: 10.1210/en.2003-0307
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Sharma, A. , Hoeffer, C.A., Takayasu, Y., Miyawaki, T., McBride, S.M., Klann, E., et al. (2010). Нарушение регуляции передачи сигналов mTOR при синдроме ломкой Х-хромосомы. J. Neurosci. 30, 694–702. doi: 10.1523/jneurosci.3696-09.2010
, Hoeffer, C.A., Takayasu, Y., Miyawaki, T., McBride, S.M., Klann, E., et al. (2010). Нарушение регуляции передачи сигналов mTOR при синдроме ломкой Х-хромосомы. J. Neurosci. 30, 694–702. doi: 10.1523/jneurosci.3696-09.2010
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Сигурдссон, Т., Старк, К.Л., Карайоргу, М., Гогос, Дж.А., и Гордон, Дж.А. (2010). Нарушение гиппокампально-префронтальной синхронии в генетической мышиной модели шизофрении. Природа 464, 763–767. doi: 10.1038/nature08855
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Судзуки, А., Stern, S.A., Bozdagi, O., Huntley, G.W., Walker, R.H., Magistretti, P.J., et al. (2011). Транспорт лактата между астроцитами и нейронами необходим для формирования долговременной памяти. Сотовый 144, 810–823. doi: 10.1016/j.cell.2011.02.018
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Танака Дж. , Хориике Ю., Мацудзаки М., Миядзаки Т., Эллис-Дэвис Г. К. и Касаи Х. (2008). Белковый синтез и нейротрофинзависимая структурная пластичность одиночных дендритных шипиков. Наука 319, 1683–1687. doi: 10.1126/science.1152864
, Хориике Ю., Мацудзаки М., Миядзаки Т., Эллис-Дэвис Г. К. и Касаи Х. (2008). Белковый синтез и нейротрофинзависимая структурная пластичность одиночных дендритных шипиков. Наука 319, 1683–1687. doi: 10.1126/science.1152864
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Танда, К., Ниши, А., Мацуо, Н., Наканиси, К., Ямасаки, Н., Сугимото, Т., и др. (2009). Аномальное социальное поведение, гиперактивность, нарушение отдаленной пространственной памяти и усиление D1-опосредованной дофаминергической передачи сигналов у мышей с нокаутом нейрональной синтазы оксида азота. Мол. Мозг 2:19. дои: 10.1186/1756-6606-2-19
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Тони, Н., Buchs, P.A., Nikonenko, I., Bron, C.R., and Muller, D. (1999). LTP способствует образованию множественных шипиковых синапсов между одним окончанием аксона и дендритом.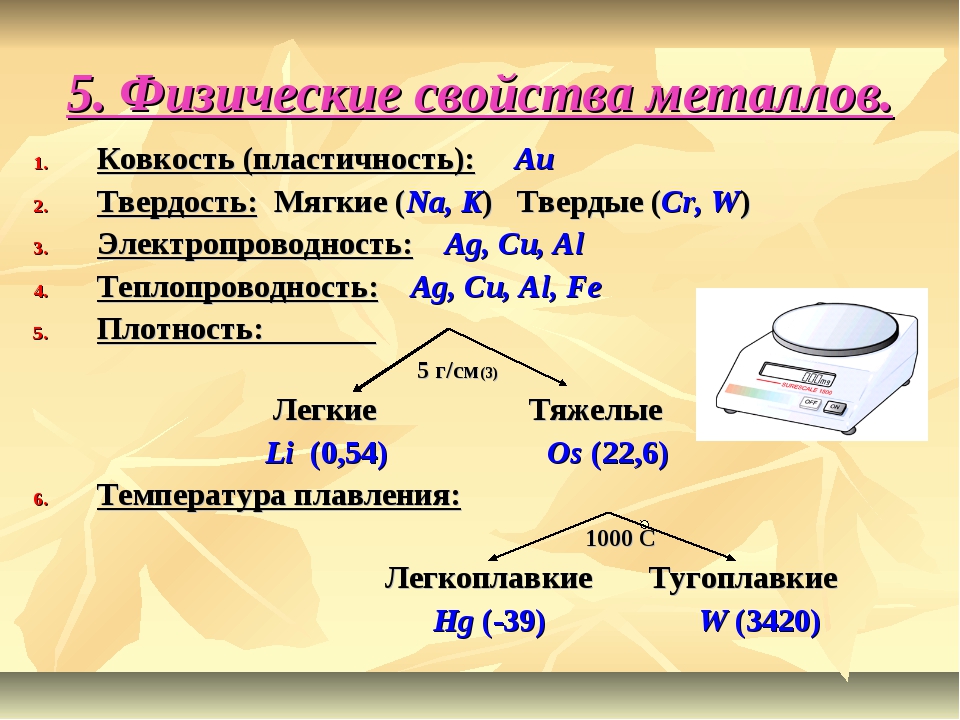 Природа 402, 421–425. дои: 10.1038/46574
Природа 402, 421–425. дои: 10.1038/46574
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Тони, Н., Тенг, Э. М., Бушонг, Э. А., Аймоне, Дж. Б., Чжао, К., Консильо, А., и соавт. (2007). Формирование синапсов на нейронах, рожденных во взрослом гиппокампе. Нац. Неврологи. 10, 727–734.дои: 10.1038/nn1908
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Вукоевич В., Гшвинд Л., Фоглер К., Демугин П., де Кервен Д. Дж., Папасситропулос А. и соавт. (2012). Роль альфа-аддуцина (ADD-1) в памяти нематод и человека. EMBO J. 31, 1453–1466. doi: 10.1038/emboj.2012.14
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Ван, Х. Б., Боздаги, О., Никитчук, Дж.С., Чжай, З.В., Чжоу, К., и Хантли, Г.В. (2008). Внеклеточный протеолиз матриксной металлопротеиназой-9 координирует увеличение дендритных шипов и долговременное потенцирование. Проц. Натл. акад. науч. США 105, 19520–19525. doi: 10.1073/pnas.0807248105
Проц. Натл. акад. науч. США 105, 19520–19525. doi: 10.1073/pnas.0807248105
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Wijetunge, L.S., Angibaud, J., Frick, A., Kind, P.C., and Nägerl, U.V. (2014). Микроскопия стимулированного эмиссионного истощения (STED) выявляет наноразмерные дефекты в траектории развития морфогенеза дендритных шипов в мышиной модели синдрома ломкой Х-хромосомы. J. Neurosci. 34, 6405–6412. doi: 10.1523/jneurosci.2974-14.2014
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Woolfrey, K.M., Srivastava, D.P., Photowala, H., Yamashita, M., Barbolina, M.V., Cahill, M.E., et al. (2009). Epac2 индуцирует ремоделирование синапсов и депрессию, а его формы, связанные с заболеванием, изменяют шипы. Нац. Неврологи. 12, 1275–1284. doi: 10.1038/nn.2386
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Сюй, Т. , Yu, X., Perlik, A.J., Tobin, W.F., Zweig, J.A., Tennant, K., et al. (2009). Быстрое формирование и избирательная стабилизация синапсов для устойчивых моторных воспоминаний. Природа 462, 915–919. doi: 10.1038/nature08389
, Yu, X., Perlik, A.J., Tobin, W.F., Zweig, J.A., Tennant, K., et al. (2009). Быстрое формирование и избирательная стабилизация синапсов для устойчивых моторных воспоминаний. Природа 462, 915–919. doi: 10.1038/nature08389
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Янкова, М., Харт, С.А., и Вулли, К.С. (2001). Эстроген увеличивает синаптическую связь между отдельными пресинаптическими входами и множественными постсинаптическими пирамидными клетками СА1: серийное электронно-микроскопическое исследование. Проц. Натл. акад. науч. США 98, 3525–3530. doi: 10.1073/pnas.051624598
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Ясумацу Н., Мацудзаки М., Миядзаки Т., Ногучи Дж. и Касаи Х. (2008). Принципы многолетней динамики дендритных шипиков. J. Neurosci. 28, 13592–13608. doi: 10.1523/jneurosci.0603-08.2008
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Зито, К. , Шойс В., Нотт Г., Хилл Т. и Свобода К. (2009). Быстрое функциональное созревание зарождающихся дендритных шипиков. Нейрон 61, 247–258. doi: 10.1016/j.neuron.2008.10.054
, Шойс В., Нотт Г., Хилл Т. и Свобода К. (2009). Быстрое функциональное созревание зарождающихся дендритных шипиков. Нейрон 61, 247–258. doi: 10.1016/j.neuron.2008.10.054
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Зубер Б., Никоненко И., Клаузер П., Мюллер Д. и Дубоше Дж. (2005). Синаптическая щель центральной нервной системы млекопитающих содержит высокую плотность периодически организованных комплексов. Проц. Натл. акад. науч. США 102, 19192–19197.doi: 10.1073/pnas.0509527102
Опубликовано Резюме | Опубликован полный текст | Полный текст перекрестной ссылки | Академия Google
Нейропластичность — StatPearls — NCBI Bookshelf
Непрерывное обучение
Нейропластичность, также известная как нейропластичность или пластичность мозга, представляет собой процесс, включающий адаптивные структурные и функциональные изменения в мозге. Он определяется как способность нервной системы изменять свою активность в ответ на внутренние или внешние раздражители путем реорганизации своей структуры, функций или связей после травм, таких как инсульт или черепно-мозговая травма (ЧМТ). В этом задании описывается нейропластичность, оценка и управление нейропластичностью, а также рассматривается роль межпрофессиональной команды в улучшении ухода за пациентами.
В этом задании описывается нейропластичность, оценка и управление нейропластичностью, а также рассматривается роль межпрофессиональной команды в улучшении ухода за пациентами.
Цели:
Обобщите рекомендации по лечению пациентов с инсультом, использующих нейропластичность.
Опишите классические результаты визуализации, связанные с диашизом.
Определите факторы риска, связанные с плохой синаптической пластичностью.
Объясните важность сотрудничества и общения внутри межпрофессиональной команды для улучшения оказания помощи пациентам с инсультом в связи с нейропластичностью.
Введение
Нейропластичность, также известная как нейронная пластичность или пластичность мозга, представляет собой процесс, включающий адаптивные структурные и функциональные изменения в головном мозге. Хорошее определение — это «способность нервной системы изменять свою активность в ответ на внутренние или внешние раздражители путем реорганизации своей структуры, функций или связей. ”[1] Клинически это процесс изменений головного мозга после травмы, такой как инсульт или черепно-мозговая травма (ЧМТ). Эти изменения могут быть как благоприятными (восстановление функции после травмы), нейтральными (без изменений), так и негативными (могут иметь патологические последствия).
”[1] Клинически это процесс изменений головного мозга после травмы, такой как инсульт или черепно-мозговая травма (ЧМТ). Эти изменения могут быть как благоприятными (восстановление функции после травмы), нейтральными (без изменений), так и негативными (могут иметь патологические последствия).
Нейропластичность можно разделить на два основных механизма:
Регенерация нейронов/коллатеральное прорастание: сюда входят такие понятия, как синаптическая пластичность и нейрогенез.
Функциональная реорганизация: сюда входят такие понятия, как эквипотенциальность, викариация и диашиз.[2] Однако термин «нейронная пластичность» приписывают Ежи Конорскому в 1948 году[1] и популяризировал Дональд Хебб в 1949 году[3].
Проблемы, вызывающие озабоченность
Пластичность после травм: Нейропластичность — сложный процесс, который все еще изучается; однако эта концепция может применяться при травме головного мозга. Традиционно считается, что нейропластичность состоит из 3 фаз или эпох.
 [4][5]
[4][5]- Первые 48 часов: В зависимости от механизма повреждения (например, инсульт или ЧМТ) начальное повреждение накапливается в виде гибели клеток с потерей определенных корковых путей, связанных с утраченными нейронами.Мозг пытается использовать вторичные нейронные сети для поддержания функции.[4]
- Следующие недели: Рекрутирование опорных клеток происходит в этот период по мере того, как корковые пути переключаются с тормозных на возбуждающие. Синаптическая пластичность и новые связи создаются в этот период.[4]
- Спустя несколько недель или месяцев: мозг продолжает перестраиваться за счет прорастания аксонов и дальнейшей реорганизации вокруг повреждения.[4]
Механизмы нейропластичности
1) Регенерация нейронов/прорастание коллатералей
Синаптическая пластичность: Синаптическая пластичность — это способность вносить зависящие от опыта долговременные изменения прочности нейронных связей.[1] Это лучше всего выражается концепцией долгосрочного потенцирования.
 Впервые обнаруженная в 1973 году Блиссом и Ломо [6] при изучении гиппокампа кролика, повторяющаяся стимуляция пресинаптических волокон приводила к высокой реакции зернистых клеток постсинаптических нейронов [7]. Поскольку постсинаптический потенциал сохранялся гораздо дольше, чем ожидалось, они назвали это долговременной потенциацией. Теоретически предполагается, что когда пресинаптический нейрон стимулирует постсинаптический нейрон, постсинаптический нейрон отвечает, добавляя больше рецепторов нейротрансмиттеров, что снижает порог, необходимый для стимуляции пресинаптического нейрона.Это усиливает синапс с течением времени в соответствии с идеей Конорски и Хебба. На синаптическую пластичность могут положительно влиять несколько вещей, включая, помимо прочего, физические упражнения, окружающую среду, повторение задач, мотивацию, нейромодуляторы (такие как дофамин) и лекарства / наркотики. ][12] Старение и нейродегенеративные заболевания связаны с уменьшением количества нейромодуляторов и могут способствовать снижению способности синаптической пластичности.
Впервые обнаруженная в 1973 году Блиссом и Ломо [6] при изучении гиппокампа кролика, повторяющаяся стимуляция пресинаптических волокон приводила к высокой реакции зернистых клеток постсинаптических нейронов [7]. Поскольку постсинаптический потенциал сохранялся гораздо дольше, чем ожидалось, они назвали это долговременной потенциацией. Теоретически предполагается, что когда пресинаптический нейрон стимулирует постсинаптический нейрон, постсинаптический нейрон отвечает, добавляя больше рецепторов нейротрансмиттеров, что снижает порог, необходимый для стимуляции пресинаптического нейрона.Это усиливает синапс с течением времени в соответствии с идеей Конорски и Хебба. На синаптическую пластичность могут положительно влиять несколько вещей, включая, помимо прочего, физические упражнения, окружающую среду, повторение задач, мотивацию, нейромодуляторы (такие как дофамин) и лекарства / наркотики. ][12] Старение и нейродегенеративные заболевания связаны с уменьшением количества нейромодуляторов и могут способствовать снижению способности синаптической пластичности. [13] Теория синаптической пластичности также расширилась, включив в себя все больше развивающейся сложности синаптической коммуникации.[1]
[13] Теория синаптической пластичности также расширилась, включив в себя все больше развивающейся сложности синаптической коммуникации.[1] К ним относятся:
Пластичность, зависящая от времени спайка (STDP): включает синхронизацию потенциалов действия, генерируемых пресинаптическими и постсинаптическими нейронами, чтобы объяснить, как одни синапсы усиливаются, а другие ослабевают.
Метапластичность. Это понятие расширяет понятие, включая сети и включает зависимые от активности изменения в синапсах и то, как они реагируют.
Гомеостатическая пластичность: механизмы, поддерживающие гомеостаз синаптической сети во времени.
По мере того, как исследования продолжают расширяться, эти концепции будут конкретизировать то, как синаптическая пластичность может влиять на обучение и помогать в восстановлении функций мозга.
Нейрогенез у взрослых. Нейрогенез у взрослых — это концепция, согласно которой мозг продолжает создавать новые нейроны.
 Исследования Рамона Кахаля не смогли найти никаких доказательств развития новых нейронов у взрослых, что привело к его «суровому указу» о том, что после остановки развития мозга новых нейронов не возникает.[14] Эта точка зрения сохранялась до тех пор, пока Джозеф Альтман не смог найти доказательства нейрогенеза у взрослых крыс.[14] С тех пор нейрогенез удалось обнаружить у птиц и других мелких млекопитающих. Это не было убедительно продемонстрировано на людях.[15]
Исследования Рамона Кахаля не смогли найти никаких доказательств развития новых нейронов у взрослых, что привело к его «суровому указу» о том, что после остановки развития мозга новых нейронов не возникает.[14] Эта точка зрения сохранялась до тех пор, пока Джозеф Альтман не смог найти доказательства нейрогенеза у взрослых крыс.[14] С тех пор нейрогенез удалось обнаружить у птиц и других мелких млекопитающих. Это не было убедительно продемонстрировано на людях.[15]Было высказано предположение о двух участках нейрогенеза у взрослых людей: одно в обонятельной луковице, а другое в гиппокампе. Исследования с использованием специфических биомаркеров, связанных с развивающимися нейронами, использовались для поддержки идеи нейрогенеза у взрослых людей. Эти биомаркеры сложны, поскольку они также были обнаружены в незрелых нейронах, клетках, которые можно найти в человеческом мозге, которые не являются новорожденными или мигрирующими клетками.[15] В сочетании с отсутствием признаков узнаваемой нишевой структуры при гистологическом исследовании (что наблюдается у других видов, проявляющих взрослый нейрогенез), доказательства неубедительны.
 Скорее всего, потребуется разработать более специфические биомаркеры для идентификации новорожденных нейронов от незрелых нейронов, чтобы выяснить, какую роль они могут играть в пластичности мозга. [15]
Скорее всего, потребуется разработать более специфические биомаркеры для идентификации новорожденных нейронов от незрелых нейронов, чтобы выяснить, какую роль они могут играть в пластичности мозга. [15]2) Функциональная реорганизация
Эквипотенциальность и викариация: Эквипотенциальность — это концепция, согласно которой при повреждении одной области мозга противоположная сторона мозга сможет поддерживать утраченную функцию.Эта концепция восходит, по крайней мере, к Галену, который был способом объяснить, почему мозг кажется «двойным». Эта «теория избыточности» существовала до тех пор, пока знаменитые исследователи, такие как Пьер Поль Брока, не продемонстрировали, что односторонние поражения в области левой стороны мозга мозг вызвал потерю речи, хотя противоположная сторона была цела. Брока постулировал, что повторное обучение определенным функциям, таким как речь, для ребенка было легче, чем если бы взрослый потерпел утрату. Эта концепция трансформировалась в эквипотенциальность, означающую, что если повреждение произошло очень рано, то у мозга есть потенциал, чтобы восполнить утраченные функции.
 [16]
[16]Это немного отличается от идеи викариации, которая заключается в том, что мозг может реорганизовать другие части мозга, чтобы взять на себя функции, для которых они не предназначены. Брока разработал эту теорию после того, как увидел, что у некоторых из его пациентов сохранялась функция, даже если у них было повреждение левого полушария. Строго говоря, викариация — это когда часть мозга берет на себя новую и несвязанную функцию. С появлением передовых методов визуализации было показано, что ни одна из теорий не совсем верна.
Гравелин, Микулис, Кроули и Хван могли показать, что после гемисферэктомии (при которой удаляется половина коры головного мозга, как правило, из-за трудноизлечимых припадков в молодом возрасте) мозг может реорганизовать оставшуюся половину, чтобы восстановить потерянная функция.[17] Используя функциональную магнитно-резонансную томографию (МРТ), они смогли показать, что оставшиеся дополнительные двигательные и сенсорные области могут быть реорганизованы, чтобы взять на себя функцию пораженной стороны.
 Джайлард и др.[18] смогли продемонстрировать аналогичные результаты у 4 взрослых пациентов, перенесших ишемический инсульт правой первичной моторной коры. Проведя серийную функциональную МРТ, они смогли показать, что мозг был способен первоначально проявлять повышенную активность в билатеральной премоторной коре, которая со временем сместилась в дополнительную моторную кору правого полушария. Эти клинические примеры подчеркивают, что мозг использует как эквипотенциальность, так и викариацию.
Джайлард и др.[18] смогли продемонстрировать аналогичные результаты у 4 взрослых пациентов, перенесших ишемический инсульт правой первичной моторной коры. Проведя серийную функциональную МРТ, они смогли показать, что мозг был способен первоначально проявлять повышенную активность в билатеральной премоторной коре, которая со временем сместилась в дополнительную моторную кору правого полушария. Эти клинические примеры подчеркивают, что мозг использует как эквипотенциальность, так и викариацию.Диасхизис: Диасхизис — это концепция, при которой повреждение одной части мозга может привести к потере функции в другой области из-за какого-то связанного пути.[19] Константин фон Монаков предложил эту концепцию в попытке объяснить, почему некоторые люди потеряли определенные функции (например, речь), но не имели поражения в области мозга, которая, как считается, обеспечивала эту функцию.[20]
Примером этого является гипоперфузия ипсилатерального таламуса после острого инсульта средней мозговой артерии (СМА).
 Таламус, который получает кровоснабжение из ветвей задней мозговой артерии и ветви задней соединительной артерии, не должен быть затронут во время ипсилатерального ишемического инсульта СМА.Удивительно, но примерно в 20% острых инсультов СМА отмечается гипоперфузия ипсилатерального таламуса при визуализации перфузии с помощью компьютерной томографии (КТ) [21]. Другие исследования показали, что заболеваемость возрастает в подострой и хронической фазах инсульта до 86%, и хотя причина до сих пор неизвестна, преобладающая теория состоит в том, что происходит растормаживание вследствие потери гамма-аминомасляной кислоты (ГАМК-энергетической) нейронов, что приводит к сочетанию нейротоксичности и ретроградной дегенерации.[21] Хотя это явление было отмечено, в настоящее время не было показано, что оно влияет на какие-либо важные клинические исходы.
Таламус, который получает кровоснабжение из ветвей задней мозговой артерии и ветви задней соединительной артерии, не должен быть затронут во время ипсилатерального ишемического инсульта СМА.Удивительно, но примерно в 20% острых инсультов СМА отмечается гипоперфузия ипсилатерального таламуса при визуализации перфузии с помощью компьютерной томографии (КТ) [21]. Другие исследования показали, что заболеваемость возрастает в подострой и хронической фазах инсульта до 86%, и хотя причина до сих пор неизвестна, преобладающая теория состоит в том, что происходит растормаживание вследствие потери гамма-аминомасляной кислоты (ГАМК-энергетической) нейронов, что приводит к сочетанию нейротоксичности и ретроградной дегенерации.[21] Хотя это явление было отмечено, в настоящее время не было показано, что оно влияет на какие-либо важные клинические исходы.Понятие диашиз со временем расширилось и используется для объяснения нескольких различных концепций о функциональных связях мозга и о том, что происходит при повреждении.
 Хотя они обсуждаются Каррерой и Тонони [19], дается краткое объяснение:
Хотя они обсуждаются Каррерой и Тонони [19], дается краткое объяснение:Диашизис «в покое»: классический тип фон Монакова, такой как ипсилатеральная таламическая гипоперфузия при инсульте СМА.
Функциональный диашиз: это когда обнаруживается область диашизиса при активации другой части мозга. Примером этого является поражение скорлупы, когда функциональная задача их ипсилатеральной руки вызывает гипоактивацию ипсилатерального мозжечка, который не имел признаков гипоактивации в состоянии покоя. Динамический диашиз также можно использовать, и он использовался, когда области мозга могут быть как гипоактивными, так и гиперактивными, в зависимости от задачи.
- Диашизис связи: это когда потеря части мозга приводит к перенаправлению информации.Это было замечено на моделях крыс, где подкорковые поражения могут вызывать снижение межполушарной связи моторных полос.[22]
Диашизис коннектома: Поскольку расширенная визуализация показала огромную сложность связей между нейронами, можно создать карту, называемую коннектомом.
 На этой карте показаны кластеры узлов с высокой степенью связи, которые затем связаны ограниченным числом узлов (концентраторов). Если повреждение нанесено концентратору, это может привести к гораздо более серьезному повреждению, чем узлу, не являющемуся концентратором.
На этой карте показаны кластеры узлов с высокой степенью связи, которые затем связаны ограниченным числом узлов (концентраторов). Если повреждение нанесено концентратору, это может привести к гораздо более серьезному повреждению, чем узлу, не являющемуся концентратором.
По мере того, как мы узнаем больше о функциональных связях мозга, концепция и роль диашизиса будут продолжать развиваться и меняться.
Клиническое значение
Нейропластичность, процесс структурных и функциональных изменений в головном мозге после внутреннего или внешнего повреждения, является общим термином, который включает множество различных процессов. Синаптическая пластичность, функциональная реорганизация и диашиз демонстрируют уникальные процессы, которые мозг использует в ответ на повреждение и восстановление функции.Поскольку исследования продолжают изучать функциональные связи в мозге и то, что влияет на эти связи, мы сможем разработать более целенаправленные методы лечения, чтобы помочь мозгу быстрее и полнее восстановить функции.

Клинически можно использовать несколько вариантов лечения, чтобы помочь направить нейропластичность в восстановлении функции и лечении нежелательных симптомов. Примером может служить зеркальная терапия, метод, используемый при фантомных болях в конечностях. В основной предпосылке патент использует зеркало, чтобы скрыть их ампутацию, и фокусируется на наблюдении за их неповрежденной конечностью, выполняющей действия, при этом представляя, что обе конечности выполняют одно и то же действие.Было показано, что это увеличивает активацию и функциональную связь в лобно-теменной сети.
Одним из наиболее изученных методов реабилитации является двигательная терапия, индуцированная ограничениями (CIMT). При использовании у пациентов с инсультом предпосылка заключается в том, что, ограничивая функциональную конечность, пораженная конечность участвует в выполнении повторяющихся задач и формировании поведения. Используя технологию функциональной магнитно-резонансной томографии (фМРТ), у пациентов, которые проходят эту терапию, было показано повышение активности в их контралатеральной премоторной и вторичной соматосенсорной коре в связи с улучшением функции.
 [23]
[23]В то время как терапия может использоваться для улучшения нейропластичности, несколько лекарств также могут использоваться для воздействия на заживление мозга. К ним относятся селективные ингибиторы обратного захвата серотонина (СИОЗС), такие как флуоксетин, ингибиторы обратного захвата серотонина и норадреналина (СИОЗСН), такие как дулоксетин, холинергические агонисты, такие как донепезил, глутаминергические частичные антагонисты, такие как амантадин, и некоторые другие [12]. Было показано, что амантадин улучшает восстановление у пациентов в минимальном сознании или в вегетативном состоянии после тяжелой ЧМТ.[24] Также было показано, что амантадин увеличивает активацию левой префронтальной коры в связи с улучшением когнитивных функций у пациентов с хронической ЧМТ.[25] По мере продолжения исследований мы сможем использовать фармакологические методы лечения, чтобы помочь мозгу вернуться к здоровому состоянию.
Неадекватная пластичность: хотя нейропластичность может быть полезной (т.
 е. восстанавливать функцию), она также может быть вредной. Дезадаптивная пластичность — это когда связь, установленная в мозгу, вызывает аберрантные или негативные симптомы.Это можно увидеть на примере дистонии, зависящей от употребления (писчий спазм), и фантомных болей в конечностях.[9] Оба этих примера показали аномальные изменения первичной сенсорной коры в сочетании с болезненными симптомами.
е. восстанавливать функцию), она также может быть вредной. Дезадаптивная пластичность — это когда связь, установленная в мозгу, вызывает аберрантные или негативные симптомы.Это можно увидеть на примере дистонии, зависящей от употребления (писчий спазм), и фантомных болей в конечностях.[9] Оба этих примера показали аномальные изменения первичной сенсорной коры в сочетании с болезненными симптомами.Наконец, большое количество исследований было сосредоточено на влиянии на нейропластичность путем модификации факторов окружающей среды. Было показано, что музыкальная терапия положительно влияет на нейропластичность. Было показано, что он улучшает познание и другие исполнительные функции. Было показано, что физические упражнения улучшают эпизодическую память и скорость обработки информации, а также уменьшают возрастную атрофию гиппокампа.[28] Здоровая диета также помогает в этом. Изучаются различные пищевые добавки, чтобы выяснить, могут ли они помочь вызвать нейропластичность. Наконец, было показано, что снижение стресса и недосыпание помогают улучшить память, концентрацию внимания и другие области познания.
 [29][30]
[29][30]Улучшение результатов медицинских бригад
Межпрофессиональный командный подход с участием неврологов, физиотерапевтов, терапевтов, медсестер и других медицинских работников, занимающихся реабилитацией пациентов после неврологических травм, обеспечит наилучшие результаты для пациентов; однако литература ограничена в оценке этого подхода к уходу.[31] Межпрофессиональная команда сможет определить наиболее подходящие варианты лечения для использования нейропластичности. Сотрудничество, совместное принятие решений и общение являются ключевыми факторами для достижения хороших результатов. Межпрофессиональная помощь, оказываемая пациенту, должна использовать комплексный подход в сочетании с доказательным подходом к планированию и оценке всех мероприятий. Несмотря на то, что телереабилитация становится все более популярной, она все еще нуждается в дополнительных исследованиях, чтобы определить ее влияние на мозг и заживление [31]. Это может помочь использовать технологии для использования телемедицины как части комплексного командного подхода к уходу за пациентами.
 [Уровень 5]
[Уровень 5]Фенотипическая пластичность растений в ответ на факторы окружающей среды
Растения подвергаются неоднородному воздействию окружающей среды, в которой появляются новые стрессовые факторы (например, изменение климата, изменения в различия могут отражать ограниченность ресурсов и/или факторы стресса окружающей среды. Фенотипическая пластичность считается одним из основных средств, с помощью которых растения могут справляться с изменчивостью факторов окружающей среды. Тем не менее, степень, в которой фенотипическая пластичность может способствовать выживанию при изменении условий окружающей среды, все еще остается в значительной степени неизвестной, поскольку результаты иногда противоречивы.Таким образом, важно определить функциональные признаки растений, пластичность которых может играть определяющую роль в реакции растений на глобальные изменения, а также на экологические последствия конкуренции между дикими и инвазивными видами на уровне экосистемы, учитывая, что виды с большей адаптивная пластичность может иметь больше шансов выжить в новых условиях окружающей среды.
 В ближайшем будущем будет важно увеличить долгосрочные исследования природных популяций, чтобы понять реакцию растений на колебания факторов окружающей среды, включая изменение климата.Существует необходимость анализа вариаций на фенотипическом и генетическом уровнях для одних и тех же видов и, в частности, для эндемичных и редких видов, поскольку они могут иметь серьезные последствия на уровне экосистемы.
В ближайшем будущем будет важно увеличить долгосрочные исследования природных популяций, чтобы понять реакцию растений на колебания факторов окружающей среды, включая изменение климата.Существует необходимость анализа вариаций на фенотипическом и генетическом уровнях для одних и тех же видов и, в частности, для эндемичных и редких видов, поскольку они могут иметь серьезные последствия на уровне экосистемы.1. Введение
Литература по фенотипической пластичности увеличилась, перейдя от первоначального внимания к абиотическим факторам к биотическим [1–3] и, в последние годы, принимая во внимание реакцию растений на глобальное изменение климата, изменения в землепользовании. инвазивность растений [4, 5] (табл. 1).Таким образом, фундаментальные вопросы для экологов-эволюционистов в контексте глобальных изменений заключаются в том, как виды растений будут реагировать на эти новые сценарии и какие механизмы будут задействованы в этом процессе [6, 7]. Понимание фенотипической пластичности будет иметь решающее значение для прогнозирования изменений в распространении видов, составе сообществ и продуктивности сельскохозяйственных культур в условиях глобальных изменений [8, 9].
 Тем не менее, тема фенотипической пластичности сложна, и исследователи не всегда приходят к одним и тем же выводам, а результаты иногда противоречивы.
Тем не менее, тема фенотипической пластичности сложна, и исследователи не всегда приходят к одним и тем же выводам, а результаты иногда противоречивы.Acer PseudoMlatanus Acer Saccharum [28]Campanula Thyrsoides [29]Coffea Arabica [30, 31 ] Corylus avellana [32–34] Crepis pygmaea subsp. PYGMAEA [35] Cyclobalanopsis MultiNervis EPILOBIUM FLOISCERI FAGU SYLVATICA [27, 37-39 Fraxinus Excelsior [32-34] 6Geum RePtans глицин Макс [40] 6глицин Soja [40] Hedera Helix 9 9ILEX Aquifolium Isatis Apennina [35]6Lythrum salicaria [43, 44] Myrtus communis [45] 90 618 Ostraa Virginiana [28]Phillyrea Latifolia [41, 46] 9 [27, 47]Picea Amies Pinus Halepensis [48-50] Pinus Nigra Pinus Picaster 5 [48] Pinus Pinea Pinus Sylvestris [48, 51] 6Pinus Uncinata Pistacia LENTISCUS 60 [46, 52-55]
POA Alpina 5 29] Populus tremula [32, 33] Populus tremuloides [28] 16Quercu s алиана вар.  ACUTISERTARATA
ACUTISERTARATA [36] Quercus Coccifera Quercus Faginea 6 [48] Quercus Ilex 5 [19, 46 , 48, 55, 57–61] Quercus mongolica var. Crispula [62, 63] Quercus Petraea Quercus Pyrenaica Quercus Robur [48] Quercus Suber Rhododendron Ponticum Rubia Peregrina 0 [41, 64]
Ruscus Aculeatus [41] Sesleria Nitida Shorea Disticha Shorea Trapezifolia [66]6Shorea Unitingniii [66] Smilax aspera [41] Stellaria Longipes [67] Taraxacum Officinale [68] Tilia Cordata [32-34]Viburnum Tinus [41 ] Фенотипическая пластичность определяется как изменение фенотипа, выраженное одним генотипом в различных условиях.
 Брэдшоу [10] признал, что фенотипическая пластичность сама по себе может находиться под генетическим контролем и, следовательно, подвергаться селективному давлению. Шайнер и Гуднайт [11] показывают, что нет оснований полагать, что отбор пластических и генетических вариаций обязательно должен быть связан. Тем не менее, популяция может реагировать на чрезвычайно изменчивую среду, становясь более пластичной и более генетически изменчивой. Литература предполагает, что фенотипическая пластичность может развиваться при наличии достаточной генетической изменчивости [12, 13] из-за генетических корреляций с другими признаками, которые находятся в процессе селекции, или генетического дрейфа [14].Поскольку фенотипическая пластичность влияет на толерантность к окружающей среде, разные пластические реакции могут способствовать различиям в диапазоне сред, в которых обитают виды [15]. В частности, окружающая среда может вызывать изменения в поведении индивидуума на морфологическом и/или физиологическом уровне [16], и такие изменения могут иметь решающее значение для выживания в гетерогенных и изменчивых условиях [17–20].
Брэдшоу [10] признал, что фенотипическая пластичность сама по себе может находиться под генетическим контролем и, следовательно, подвергаться селективному давлению. Шайнер и Гуднайт [11] показывают, что нет оснований полагать, что отбор пластических и генетических вариаций обязательно должен быть связан. Тем не менее, популяция может реагировать на чрезвычайно изменчивую среду, становясь более пластичной и более генетически изменчивой. Литература предполагает, что фенотипическая пластичность может развиваться при наличии достаточной генетической изменчивости [12, 13] из-за генетических корреляций с другими признаками, которые находятся в процессе селекции, или генетического дрейфа [14].Поскольку фенотипическая пластичность влияет на толерантность к окружающей среде, разные пластические реакции могут способствовать различиям в диапазоне сред, в которых обитают виды [15]. В частности, окружающая среда может вызывать изменения в поведении индивидуума на морфологическом и/или физиологическом уровне [16], и такие изменения могут иметь решающее значение для выживания в гетерогенных и изменчивых условиях [17–20]. Для некоторых морфологических признаков показано, что фенотипическая пластичность относительно хорошо отражает генетические корреляции, а признаки, принадлежащие к одному и тому же набору признаков, более сильно генетически и фенотипически коррелированы, чем признаки из разных наборов [21].Отбор фотосинтетических признаков часто может осуществляться косвенно через корреляцию с другими признаками, что подчеркивает важность рассмотрения фенотипа как интегрированной функции роста, морфологии, жизненного цикла и физиологии [22]. Сроки развития растений сами по себе могут быть пластичными [23], и многие фенотипические реакции на стрессовые факторы окружающей среды могут быть следствием снижения роста из-за ограниченности ресурсов [24, 25]. Различия между видами и популяциями могут отражать различное селективное давление на пластичность, различные ограничения, действующие на максимизацию пластичности, или их комбинацию [26].Потенциальная пластическая реакция данного признака может быть большой, но наблюдаемая пластичность может быть снижена из-за ограниченности ресурсов или факторов стресса окружающей среды [14].
Для некоторых морфологических признаков показано, что фенотипическая пластичность относительно хорошо отражает генетические корреляции, а признаки, принадлежащие к одному и тому же набору признаков, более сильно генетически и фенотипически коррелированы, чем признаки из разных наборов [21].Отбор фотосинтетических признаков часто может осуществляться косвенно через корреляцию с другими признаками, что подчеркивает важность рассмотрения фенотипа как интегрированной функции роста, морфологии, жизненного цикла и физиологии [22]. Сроки развития растений сами по себе могут быть пластичными [23], и многие фенотипические реакции на стрессовые факторы окружающей среды могут быть следствием снижения роста из-за ограниченности ресурсов [24, 25]. Различия между видами и популяциями могут отражать различное селективное давление на пластичность, различные ограничения, действующие на максимизацию пластичности, или их комбинацию [26].Потенциальная пластическая реакция данного признака может быть большой, но наблюдаемая пластичность может быть снижена из-за ограниченности ресурсов или факторов стресса окружающей среды [14].
Конкретный способ, которым генотип варьирует в своей экспрессии в различных средах, может быть описан нормой реакции, которая определяется генетически [69]. Норма реакции на любой конкретный признак генотипа может быть визуализирована в виде линии или кривой на двумерном графике зависимости средовой ценности от фенотипической (рис. 1).Фенотипическую пластичность можно представить как изменение наклона нормы реакции между предковыми и производными популяциями или видами [70, 71]. Было показано, что такие изменения происходят в природе между видами, подвергающимися различному давлению отбора [72–74]. Пластичность — это то, что делает возможным появление индуцированного средой нового фенотипа, и процесс отбора на выражение такого фенотипа в новой среде может закончиться «закреплением» (генетической ассимиляцией) его путем изменения формы нормы реакции [75]. ].Таким образом, пластичность может способствовать выражению относительно хорошо адаптированных фенотипов в новых условиях (например, после миграции в новые географические районы), повышая производительность популяции и приводя к генетической ассимиляции признака в новой среде.
 Это может объяснить множество эволюционных экологических процессов [14, 75].
Это может объяснить множество эволюционных экологических процессов [14, 75].
Индексы могут облегчить сравнение различных исследований [76, 77], наборов видов или популяций внутри данного вида с учетом экспериментальных данных в исследованиях пластичности [19, 56, 57].Однако в качестве меры фенотипической пластичности использовалось по крайней мере 17 различных индексов, но они могут иметь недостатки и применяться по-разному. Большинство из них нельзя стандартизировать по признакам или сравнивать между разными видами [78], что усложняет сравнение между исследованиями [3, 79]. Более того, показатели фенотипической пластичности тесно связаны с контекстом и могут быть несопоставимы в разных исследованиях, в которых использовались разные градиенты и/или виды [77]. Поскольку информация о пластичности структурирована таким образом, что ее количественный и сравнительный анализ затруднен, Poorter et al.[79] предложили метод заполнения этого пробела путем создания большой базы данных, которая в настоящее время содержит данные о 1000 экспериментах и 800 видах.
 Этот подход может служить ориентиром для будущих усилий по фенотипированию, а также для моделирования последствий глобальных изменений как для диких видов, так и для сельскохозяйственных культур [79].
Этот подход может служить ориентиром для будущих усилий по фенотипированию, а также для моделирования последствий глобальных изменений как для диких видов, так и для сельскохозяйственных культур [79].2. Физиологическая, анатомическая и морфологическая пластичность
Физиологическая, морфологическая и анатомическая пластичность могут играть различную роль в приспособлении растений к изменениям среды.В частности, пластичность физиологических и жизненных признаков может позволить растениям расти и размножаться в пространственно или во времени изменчивых условиях [46, 80]. Физиологическая пластичность в большей степени связана с повышенной способностью заселять щели и открытые пространства [81, 82], так как обеспечивает перестройку газообмена в ответ на изменение факторов внешней среды в короткие сроки. Этот аспект свидетельствует о важности физиологической пластичности при адаптации растений к неблагоприятным условиям среды, где морфологическая и анатомическая пластичность играют второстепенную роль [83].
 На самом деле, растения, растущие в стрессовых условиях, как правило, имеют консервативную морфологию листьев, чтобы избежать производства структур, слишком дорогих для поддержания [84, 85]. Кроме того, морфологическая пластичность больше связана с повышенной способностью растений расти в подлеске [37, 82], играя важную роль в приобретении ресурсов [64, 86].
На самом деле, растения, растущие в стрессовых условиях, как правило, имеют консервативную морфологию листьев, чтобы избежать производства структур, слишком дорогих для поддержания [84, 85]. Кроме того, морфологическая пластичность больше связана с повышенной способностью растений расти в подлеске [37, 82], играя важную роль в приобретении ресурсов [64, 86].3. Реакция растений на свет
Неоднородная световая среда в кроне растений вызывает различные факторы стресса для листьев в разных положениях кроны.Одним из проявлений фенотипической пластичности растений является модификация признаков листа в зависимости от градиента света [87] и пониженного отношения красного/дальнего красного [2] от вершины к низу кроны дерева, в основном во время листообразования [69]. . Одним из основных морфологических признаков, изменяющихся в ответ на изменение освещенности, является удельная площадь листа (SLA, отношение площади листа к сухой массе листа). Пластичность SLA подразумевает морфогенетический контроль листьев, который имеет тенденцию к увеличению площади листьев в тени, чтобы перехватить больше света, в то время как, напротив, существует генетическое или физиологическое ограничение общего объема листьев, а также ограничение ресурсов [88].
 .SLA отражает толщину листа [89] и относительную долю ассимиляционных, проводящих и механических тканей [90]. В частности, увеличенная общая толщина пластинки солнечных листьев по сравнению с теневыми листьями в основном связана с большей палисадной паренхимой, губчатой паренхимой и толщиной эпидермальной ткани, что позволяет предположить, что внутренняя структура листа может играть важную роль в захвате света [91]. В исследовании морфологии листа древесных покрытосеменных Вайли [92] показывает, что в результате затенения толщина листа уменьшилась в среднем на 54 %, объем палисадной паренхимы — на 60 %, а толщина эпидермиса — на 17%.McClendon [93] и Gratani et al. [46] показали сильную корреляцию между толщиной листа и светонасыщенной скоростью фотосинтеза на единицу площади листа.
.SLA отражает толщину листа [89] и относительную долю ассимиляционных, проводящих и механических тканей [90]. В частности, увеличенная общая толщина пластинки солнечных листьев по сравнению с теневыми листьями в основном связана с большей палисадной паренхимой, губчатой паренхимой и толщиной эпидермальной ткани, что позволяет предположить, что внутренняя структура листа может играть важную роль в захвате света [91]. В исследовании морфологии листа древесных покрытосеменных Вайли [92] показывает, что в результате затенения толщина листа уменьшилась в среднем на 54 %, объем палисадной паренхимы — на 60 %, а толщина эпидермиса — на 17%.McClendon [93] и Gratani et al. [46] показали сильную корреляцию между толщиной листа и светонасыщенной скоростью фотосинтеза на единицу площади листа.Солнечные листья по сравнению с теневыми листьями в среднем имеют более высокую скорость фотосинтеза на основе площади листа, что связано с более высоким содержанием хлорофилла (Хл) a / b соотношение, значительно более низким светособирающим Хл a / b белок (LHCII), более низкая степень укладки тилакоидов [94, 95] и более высокое содержание азота (N) на единицу площади листа [46, 96, 97], так как примерно половина N приходится на инвестируются в фотосинтетические белки [96].
 Hirose и Werger [98] предполагают, что азот изменяется в зависимости от наличия света в растительном покрове таким образом, чтобы оптимизировать ежедневный фотосинтез в растительном покрове. Таким образом, вложение азота связано со светом в том смысле, что больше азота выделяется на карбоксилирование в солнечных листьях и на сбор света в тенистых листьях с изменением фотосинтетической способности [99] (рис. 2). Фотосинтетическая способность и содержание N, как правило, выше в листьях в условиях высокой освещенности [96, 98, 100]. Более высокая скорость фотосинтеза солнечных листьев поддерживается более высокой устьичной проводимостью и плотностью устьиц, чтобы максимизировать поглощение CO 2 [101].Напротив, приспособление листьев к слабому освещению увеличивает способность поглощения света за счет фотосинтетической способности, сводящей к минимуму потерю углерода через дыхание [96].
Hirose и Werger [98] предполагают, что азот изменяется в зависимости от наличия света в растительном покрове таким образом, чтобы оптимизировать ежедневный фотосинтез в растительном покрове. Таким образом, вложение азота связано со светом в том смысле, что больше азота выделяется на карбоксилирование в солнечных листьях и на сбор света в тенистых листьях с изменением фотосинтетической способности [99] (рис. 2). Фотосинтетическая способность и содержание N, как правило, выше в листьях в условиях высокой освещенности [96, 98, 100]. Более высокая скорость фотосинтеза солнечных листьев поддерживается более высокой устьичной проводимостью и плотностью устьиц, чтобы максимизировать поглощение CO 2 [101].Напротив, приспособление листьев к слабому освещению увеличивает способность поглощения света за счет фотосинтетической способности, сводящей к минимуму потерю углерода через дыхание [96].
Изменения в эффективности улавливания света и затратах на сбор света вдоль градиента света от вершины к низу растительного покрова являются основными средствами, с помощью которых достигается эффективный сбор света.
 В частности, в масштабе побегов угол наклона листвы и пространственное скопление листвы являются основными детерминантами сбора света, в то время как в масштабе кроны частота ветвления распределение листвы и распределение биомассы по листьям значительно изменяют сбор света [102].Пластичность внутри полога оказывает важное влияние на многие аспекты биологии дерева, потенциально способствуя производительности всего полога за счет эффектов проникновения света через полог и на энергетический, углеродный и водный баланс отдельных листьев [97, 103, 104]. Пластичность в пределах одного и того же растения во временной гетерогенной среде может играть адаптивную роль в сильном сезонном климате, таком как средиземноморский климат [20].
В частности, в масштабе побегов угол наклона листвы и пространственное скопление листвы являются основными детерминантами сбора света, в то время как в масштабе кроны частота ветвления распределение листвы и распределение биомассы по листьям значительно изменяют сбор света [102].Пластичность внутри полога оказывает важное влияние на многие аспекты биологии дерева, потенциально способствуя производительности всего полога за счет эффектов проникновения света через полог и на энергетический, углеродный и водный баланс отдельных листьев [97, 103, 104]. Пластичность в пределах одного и того же растения во временной гетерогенной среде может играть адаптивную роль в сильном сезонном климате, таком как средиземноморский климат [20].Кроме того, вариации признаков листьев в зависимости от градиента освещенности в пологе меняются у разных видов и типов леса [105–108].В лесах с густой листвой верхние слои поглощают большую часть поступающей радиации. В широколиственных и хвойных лесах умеренного пояса в крону деревьев проникает в среднем 3–10% плотности потока падающих фотонов (ППП) [69].
 Гратани и Фоти [109] показывают, что SLA увеличивается в среднем на 21% от преобладающего древесного яруса смешанного лиственно-широколиственного леса к преобладающему ярусу в ответ на снижение уровня освещенности. Та же тенденция измеряется от преобладающего слоя деревьев средиземноморского вечнозеленого леса к преобладающему слою увеличением SLA на 54%, что связано с 9.На 5% ниже соотношение хлорофилла a / b и на 86% ниже скорость фотосинтеза [110]. Мендес и др. [45] проанализировали значительные вариации морфологических и анатомических признаков листа в зависимости от условий освещения для Myrtus communis , вечнозеленого склерофильного вида, который растет в средиземноморских районах от открытых полян до подлеска, с 26-процентным увеличением SLA от солнца к тени. . Тем не менее, таксономически разные виды, сосуществующие в одной и той же среде обитания, часто имеют общие морфологические и физиологические черты, отражающие конвергентную эволюцию в ответ на факторы окружающей среды [111].
Гратани и Фоти [109] показывают, что SLA увеличивается в среднем на 21% от преобладающего древесного яруса смешанного лиственно-широколиственного леса к преобладающему ярусу в ответ на снижение уровня освещенности. Та же тенденция измеряется от преобладающего слоя деревьев средиземноморского вечнозеленого леса к преобладающему слою увеличением SLA на 54%, что связано с 9.На 5% ниже соотношение хлорофилла a / b и на 86% ниже скорость фотосинтеза [110]. Мендес и др. [45] проанализировали значительные вариации морфологических и анатомических признаков листа в зависимости от условий освещения для Myrtus communis , вечнозеленого склерофильного вида, который растет в средиземноморских районах от открытых полян до подлеска, с 26-процентным увеличением SLA от солнца к тени. . Тем не менее, таксономически разные виды, сосуществующие в одной и той же среде обитания, часто имеют общие морфологические и физиологические черты, отражающие конвергентную эволюцию в ответ на факторы окружающей среды [111]. Большая пластичность в структуре мезофилла в сочетании с другими признаками, по-видимому, позволяет этим видам справляться с различными экологическими режимами (например, средиземноморским климатом), достигая более широкого диапазона местообитаний [111].
Большая пластичность в структуре мезофилла в сочетании с другими признаками, по-видимому, позволяет этим видам справляться с различными экологическими режимами (например, средиземноморским климатом), достигая более широкого диапазона местообитаний [111].4. Теневыносливость
Тенеустойчивость является одним из важнейших экологических факторов в отношении межвидовой конкуренции среди лесных деревьев умеренного климатического пояса. Теневыносливость данного растения определяется как минимальное количество света, при котором растение может выжить; однако лишь часть растений может воспроизводиться в этих условиях.Таким образом, биологическое определение теневыносливости должно учитывать весь жизненный цикл растения от раннего выживания и роста до размножения [112]. В целом, у устойчивых к солнцу и тени листьев листья тоньше, чем у нетерпимых [113]. Общая тенденция в литературе свидетельствует о том, что раннесукцессионные и светолюбивые виды более пластичны, чем позднесукцессионные и теневыносливые виды [114–116].
 Тем не менее появляется все больше свидетельств того, что корректировки не обязательно связаны с сукцессионным статусом вида [117, 118] (табл. 2).
Тем не менее появляется все больше свидетельств того, что корректировки не обязательно связаны с сукцессионным статусом вида [117, 118] (табл. 2).90 610 Виды Морфологического пластичность Физиологическая пластичность Списка литературы + [37]6теневыносливые бук европейский 0,46 0,39 [37] Acer PseudoMlatanus 0.31 0.34 [27] ILEX Aquifolium 0.54 0.24 [119] Quercus ILEX 0.  33
330.5 ] Acer Saccharum 0.3 0.3Промежуточные Acer Rubrum 0,21 0,26 Betula Alleghaniensisis [108]0.29 0.32 [108] 9061 Picea Abies 0.13 0.22 [27]Quercus RUBRA 2020 0.41 0.27оттенок-нетерпимый Quercus Robur 0,24 0,6 Cistus Incanus 0.  18
180.38 . виды растений к разным световым условиям; тем не менее, результаты иногда расходятся. Например, Coffea arabica — вечнозеленое многолетнее дерево из подлеска африканских лесов, считающееся обязательно затеняющим видом.Матос и др. [30] показывают, что пластичность физиологических и биохимических признаков у C. arabica более важна для акклиматизации к внутрипокровным изменениям освещенности, чем пластичность морфологических и анатомических признаков. Тем не менее, Араужо и соавт. [31] показывают, что очевидная неспособность затененных листьев C. arabica оптимизировать накопление углерода при слабом освещении и успешная способность солнечных листьев предотвращать фотоингибирование фотосинтеза при ярком освещении согласуются с выводами о том, что он хорошо работает при полном освещении. воздействие солнца.Ниинеметс и др. [32] показали, что в лиственных лесах умеренного пояса виды различаются по теневыносливости: наибольшей толерантностью обладает Corylus avellana , за ним следуют Tilia cordata , Fraxinus excelsior и Populus tremula [33, 34].  Вика и др. [27] путем сравнения сеянцев двух хвойных (теневыносливого Abies alba и умеренно теневыносливого Picea abies ) и двух листопадных покрытосеменных растений (высокотеневыносливого Fagus sylvatica и умеренно теневыносливого ). Acer pseudoplatanus ) в двухгодичных исследованиях проверяли гипотезу о том, что светоиндуцированная пластичность физиологических и анатомических признаков у высокотеневыносливых видов ниже, чем у среднетеневыносливых.Авторы пришли к выводу, что теневыносливость не была постоянным предиктором анатомической пластичности и что для многих признаков различия между световыми обработками усиливались на втором году жизни, показывая, что анатомическая и физиологическая адаптация к оттенку является долгосрочным процессом. Валладарес и др. [37] исследовали фенотипическую пластичность двух видов, различающихся по теневыносливости (наиболее теневыносливого Fagus sylvatica и более светолюбивого Quercus robur ), выдвинув гипотезу о меньшей пластичности у видов глубокого затенения, чем у более светлых.
Вика и др. [27] путем сравнения сеянцев двух хвойных (теневыносливого Abies alba и умеренно теневыносливого Picea abies ) и двух листопадных покрытосеменных растений (высокотеневыносливого Fagus sylvatica и умеренно теневыносливого ). Acer pseudoplatanus ) в двухгодичных исследованиях проверяли гипотезу о том, что светоиндуцированная пластичность физиологических и анатомических признаков у высокотеневыносливых видов ниже, чем у среднетеневыносливых.Авторы пришли к выводу, что теневыносливость не была постоянным предиктором анатомической пластичности и что для многих признаков различия между световыми обработками усиливались на втором году жизни, показывая, что анатомическая и физиологическая адаптация к оттенку является долгосрочным процессом. Валладарес и др. [37] исследовали фенотипическую пластичность двух видов, различающихся по теневыносливости (наиболее теневыносливого Fagus sylvatica и более светолюбивого Quercus robur ), выдвинув гипотезу о меньшей пластичности у видов глубокого затенения, чем у более светлых. -требовательный вид, по данным сравнительных исследований других древесных растений [56, 85, 105].Более того, авторы предположили, что возможна и альтернативная гипотеза (т. е. теневыносливость может быть достигнута за счет повышенной пластичности), как утверждалось ранее для тех же двух видов [121]. Результаты показали, что средняя фенотипическая пластичность аналогична двум видам ( Q. robur и F. sylvatica ) из-за того, что в то время как Q. robur имеет более высокую физиологическую пластичность, F. sylvatica имеет более высокую морфологическая пластичность, при этом одна группа признаков противодействует влиянию другой группы на средний показатель пластичности [37].
-требовательный вид, по данным сравнительных исследований других древесных растений [56, 85, 105].Более того, авторы предположили, что возможна и альтернативная гипотеза (т. е. теневыносливость может быть достигнута за счет повышенной пластичности), как утверждалось ранее для тех же двух видов [121]. Результаты показали, что средняя фенотипическая пластичность аналогична двум видам ( Q. robur и F. sylvatica ) из-за того, что в то время как Q. robur имеет более высокую физиологическую пластичность, F. sylvatica имеет более высокую морфологическая пластичность, при этом одна группа признаков противодействует влиянию другой группы на средний показатель пластичности [37].Различия в структуре леса определяют изменения градиента освещенности и, следовательно, наличие теневыносливых и непереносимых пород. В частности, лиственные леса могут состоять из верхнего яруса с теневыносливыми древесными породами и нижнего яруса с теневыносливыми древесными породами [32], а в смешанных лесах листопадные и вечнозеленые породы с контрастным экологическим потенциалом могут занимать разные ярусы.
 122]. Путер и др. [48] проанализировали теневыносливость видов деревьев, произрастающих в некоторых лесах континентальной Испании, и показали, что более широкий полог, меньшая высота и плоские листья у Fagaceae ( Quercus faginea , Quercus ilex , Quercus petraea , Quercus pyrenaica , quercus robur и quercus suber ) Усилил их оттенок по сравнению с Pinaceae ( Pinus Halepensis , Pinus Nigra , Pinus Picaster , Pinus Pinea , Pinus Sylvestris и Pinus Uncinata ).Напротив, огнестойкость [123], засухоустойчивость [124] и аллометрическая пластичность теневыносливых сосен способствовали их быстрому заселению открытых пространств [125] и могли иметь значение для объяснения экологического доминирования видов. Теневыносливые Populus tremuloides и Betula papyrifera , распространенные виды широкого экотона между бореальными и умеренно-холодными лесами центральной и восточной Северной Америки, обладают чертами, которые максимизируют обмен двуокиси углерода (CO 2 ) всего растения.
122]. Путер и др. [48] проанализировали теневыносливость видов деревьев, произрастающих в некоторых лесах континентальной Испании, и показали, что более широкий полог, меньшая высота и плоские листья у Fagaceae ( Quercus faginea , Quercus ilex , Quercus petraea , Quercus pyrenaica , quercus robur и quercus suber ) Усилил их оттенок по сравнению с Pinaceae ( Pinus Halepensis , Pinus Nigra , Pinus Picaster , Pinus Pinea , Pinus Sylvestris и Pinus Uncinata ).Напротив, огнестойкость [123], засухоустойчивость [124] и аллометрическая пластичность теневыносливых сосен способствовали их быстрому заселению открытых пространств [125] и могли иметь значение для объяснения экологического доминирования видов. Теневыносливые Populus tremuloides и Betula papyrifera , распространенные виды широкого экотона между бореальными и умеренно-холодными лесами центральной и восточной Северной Америки, обладают чертами, которые максимизируют обмен двуокиси углерода (CO 2 ) всего растения. что приводит к проблемам длительного роста и выживания при очень слабом освещении, в то время как теневыносливые сорта Acer saccharum и Ostrya virginiana сводят к минимуму потери CO 2 при слабом освещении за счет максимизации прироста CO 2 при ярком освещении. [28].Те же авторы заключают, что теневыносливые виды обладают большей пластичностью, чем теневыносливые. Ке и Вергер [36] показали, что листопадный Quercus Aliena var. acutiserrata – щелевой вид, имеющий преимущество перед вечнозеленым Cyclobalanopsis multinervis на лесных полянах и полянах большей морфологической и физиологической пластичностью сеянцев. Способность акклиматизироваться к контрастной световой среде особенно важна для тропических древесных видов, поскольку сеянцы деревьев, выращенные в подлеске, подвержены фотоингибированию при повышенном уровне освещенности за счет образования просвета [126].Высокая освещенность в тропических широтах может вызывать хроническое фотоингибирование за счет нарушения работы реакционных центров фотосистемы II (ФСII) в листьях растений, в которых наблюдались щелевые отверстия [127, 128].
что приводит к проблемам длительного роста и выживания при очень слабом освещении, в то время как теневыносливые сорта Acer saccharum и Ostrya virginiana сводят к минимуму потери CO 2 при слабом освещении за счет максимизации прироста CO 2 при ярком освещении. [28].Те же авторы заключают, что теневыносливые виды обладают большей пластичностью, чем теневыносливые. Ке и Вергер [36] показали, что листопадный Quercus Aliena var. acutiserrata – щелевой вид, имеющий преимущество перед вечнозеленым Cyclobalanopsis multinervis на лесных полянах и полянах большей морфологической и физиологической пластичностью сеянцев. Способность акклиматизироваться к контрастной световой среде особенно важна для тропических древесных видов, поскольку сеянцы деревьев, выращенные в подлеске, подвержены фотоингибированию при повышенном уровне освещенности за счет образования просвета [126].Высокая освещенность в тропических широтах может вызывать хроническое фотоингибирование за счет нарушения работы реакционных центров фотосистемы II (ФСII) в листьях растений, в которых наблюдались щелевые отверстия [127, 128]. Тем не менее виды различаются по своей теневыносливости к повышенному освещению после создания просвета. Например, в широколиственных лесах северной Японии Quercus mongolica var. crispula является зависимым от пробелов видом, потому что его проростки могут прожить не более десятка лет под закрытым пологом, даже если они могут прорастать в условиях низкой освещенности [62, 63].Койке и др. [129] в исследовании ранних сукцессионных видов, видов в фазе разрыва, среднесукцессионных и позднесукцессионных видов показали, что потенциал акклиматизации видов выражался в их пластичности, которая была тесно связана с фотосинтетической способностью [130].
Тем не менее виды различаются по своей теневыносливости к повышенному освещению после создания просвета. Например, в широколиственных лесах северной Японии Quercus mongolica var. crispula является зависимым от пробелов видом, потому что его проростки могут прожить не более десятка лет под закрытым пологом, даже если они могут прорастать в условиях низкой освещенности [62, 63].Койке и др. [129] в исследовании ранних сукцессионных видов, видов в фазе разрыва, среднесукцессионных и позднесукцессионных видов показали, что потенциал акклиматизации видов выражался в их пластичности, которая была тесно связана с фотосинтетической способностью [130].Несмотря на предположение, что теневые листья развиваются в ответ на снижение освещенности, могут быть задействованы и другие факторы, такие как температура и водный стресс [113]. Было высказано предположение, что растения не могут переносить сочетание тени и засухи в результате морфологического компромисса [49, 131].Тем не менее, Сак и соавт.
 [41] Исследован шесть видов ( Phillyrea Latifolia , Viburnum Tinus , Rubia Peregrina , Ruscus Aculeatus , Hedera Helix , Hedera Helix , и Smilax Aspera ), которые справится с сильным летним засухой в подразделении смешанного Quercus леса на юге Испании. Все виды выживали в тени (около 3% дневного света) и сходились по признакам, которые придавали устойчивость к тени и засухе за счет снижения потребности в ресурсах.В частности, потребность в воде была снижена за счет содержания подземной массы от умеренного до высокого и SLA от низкого до среднего, в то время как потребность как в низкой освещенности, так и в воде была снижена за счет содержания азота в листьях от низкого до среднего и листьев. с длительным сроком службы. В среднем SLA является признаком, наиболее сильно коррелирующим с теневыносливостью [132].
[41] Исследован шесть видов ( Phillyrea Latifolia , Viburnum Tinus , Rubia Peregrina , Ruscus Aculeatus , Hedera Helix , Hedera Helix , и Smilax Aspera ), которые справится с сильным летним засухой в подразделении смешанного Quercus леса на юге Испании. Все виды выживали в тени (около 3% дневного света) и сходились по признакам, которые придавали устойчивость к тени и засухе за счет снижения потребности в ресурсах.В частности, потребность в воде была снижена за счет содержания подземной массы от умеренного до высокого и SLA от низкого до среднего, в то время как потребность как в низкой освещенности, так и в воде была снижена за счет содержания азота в листьях от низкого до среднего и листьев. с длительным сроком службы. В среднем SLA является признаком, наиболее сильно коррелирующим с теневыносливостью [132].Виды-универсалы, произрастающие в различных условиях влажности и освещенности в лесу, в среднем обладают большей морфологической пластичностью, чем виды-специалисты [66].В частности, более универсальная Shorea disticha , встречающаяся на большинстве участков в пределах вечнозеленых лесов и распространяющаяся на большинство районов Южной и Юго-Восточной Азии, обладает большей пластичностью, чем Shorea trapezifolia , которая ограничена нижними склонами долины в пределах лес и Shoreaworthingtonii , который ограничен вершинами хребтов.
5. Экотипы
Многолетний отбор может привести к развитию морфологических и физиологических адаптаций к локальной среде, порождающих экотипическую дифференциацию функциональных признаков [133, 134].Генотипы, адаптированные к местным условиям среды, называются экотипами [135]. Когда среды в пределах ареала распространения вида различаются, маловероятно, что какой-либо отдельный фенотип обеспечивает высокую приспособленность во всех ситуациях. Различие между фенотипической пластичностью и локальной адаптацией экотипа основано прежде всего на генетическом анализе и трансплантационных экспериментах [52]. В частности, для многих видов [136–138], а также для экотипических формаций [52, 136, 138] зафиксирована пространственная генетическая дифференциация вдоль климатических градиентов.Например, адаптивными признаками обладают экотипы Pinus taeda [139], Picea abies [47], Pinus sylvestris [51], Fagus sylvatica [38, 39], Quercus coccifera [50]. которые, вероятно, обусловлены климатом местности, из которой они происходят. Более того, виды с обширным географическим ареалом потенциально могут демонстрировать большую внутривидовую изменчивость в физиологии, морфологии, фенологии и скорости роста [140]. Гратани и др.[19], Бонито и соавт. [57] и Pesoli et al. [58] сравнили пластичность признаков растений и листьев у сеянцев Quercus ilex из разных мест происхождения в Италии и выращенных в обычном саду (рис. 3). Quercus ilex — вечнозеленый вид с глубокими корнями, широко распространенный в Средиземноморском бассейне [59] и простирающийся на 6000 км в продольном направлении от Марокко до Франции. Этот вид, по-видимому, ограничен южным ареалом из-за усиления летней засухи, а по высоте (распространен от уровня моря до 1100 м над ур.с.л.) факторами, связанными с низкой температурой [60]. Результаты показывают, что сеянцы более ксерического происхождения обладают более высокой устойчивостью к стрессу засухи за счет более высокой площади листовой массы (LMA), ограничивающей транспирацию, и более высокой чувствительности устьиц к изменениям водного потенциала, что обеспечивает более высокое относительное содержание воды в листьях (RWC). Наоборот, уменьшенная площадь листьев, по-видимому, является лучшим адаптивным признаком в ответ на зимние холода на северной границе ареала, в то время как наибольшая продукция побегов отражает более благоприятные климатические условия в центре ареала.Таким образом, в то время как благоприятные условия внешней среды повышают фенотипическую пластичность морфологических и физиологических признаков Q. ilex , менее благоприятные условия (т. е. холодовой и засушливый стресс) допускают специализацию. Мишо и др. [61] наблюдали однородную генетическую структуру Q. ilex в Средиземноморском регионе с небольшими географическими вариациями из-за изоляции (т. е. Северная Африка и Сицилия), что подтверждает гипотезу о том, что Q. ilex соответствует единому генетическому объекту. .Согласно этим гипотезам, Balaguer et al. [56] показывают, что фенотипическая пластичность варьируется в пределах географического ареала Quercus coccifera и среди популяций, предполагая, что экотипическая дифференциация Q. coccifera объясняет его появление в контрастных местообитаниях. Вен и др. [40] изучали происхождение и эволюцию культурной сои. Они исследовали генетическое разнообразие, географическую дифференциацию и генетическое родство между географическими экотипами культивируемых ( Glycine max ) и диких ( G.soja ) соевые бобы, произрастающие в Южно-Центральном Китае, Юго-Западном Китае и Южном Китае. Результаты показали, что дикие образцы имели относительно небольшие генетические расстояния со всеми культивируемыми образцами, а дикие экотипы долин Среднего и Нижнего Чанцзяна были меньше по сравнению с другими дикими образцами, включая их местные дикие аналоги. Следовательно, делается вывод, что дикие предки южного Китая, особенно из долин Среднего и Нижнего Чанцзяна, могут быть общими предками всех культивируемых соевых бобов.Тем не менее эксплуатация новых местообитаний может быть связана с утратой пластичности и эволюцией специализации [16]. Адаптация видов к географическим вариациям окружающей среды часто зависит от генетических вариаций источников семян [19, 141]. Наум и др. [52] показали, что экотипов Pistacia lentiscus , произрастающих в различных местообитаниях вдоль климатического градиента в Израиле, не имеют какой-либо закономерности экологически связанной генетической дифференциации, а морфологические и физиологические различия, вероятно, обусловлены фенотипической пластичностью.Таким образом, адаптивная пластичность может повысить устойчивость к окружающей среде, способствуя широкому распространению [53] P. lentiscus в Средиземноморском регионе [52, 54]. Эмери и др. [67] показывают, что экотипы Stellaria longipes , травянистого многолетнего вида, произрастающего вдоль высотного градиента на горе Плато (Альберта) из альпийской тундры (т.е. с большей высоты), обладают меньшей пластичностью, чем экотип из прерий (т.е. , меньшая высота).
Для точного определения паттернов пластичности и изучения экологических и эволюционных последствий важно лучше понять экологический контекст, в котором проявляются фенотипы [2].
6. Высокогорные растения
Было показано, что воздействие глобального потепления на наземные экосистемы больше в арктической тундре и высокогорных районах, чем в низких широтах [143]. В частности, ожидается, что прогнозируемая скорость глобального потепления в горных экосистемах будет в три раза выше, чем глобальная средняя скорость потепления, зарегистрированная в ХХ веке [144]. Сценарии биоразнообразия на 21 век прогнозируют сокращение альпийской среды обитания и утрату многих европейских высокогорных растений [145, 146].Дирнбёк и др. [147] выдвигают гипотезу о быстром увеличении риска исчезновения видов растений. Более того, видовое снижение приспособленности и разнообразия может изменить динамику сообщества, изменив конкурентоспособность видов. Недавние исследования [35] указывают на значительный адаптивный потенциал, о чем свидетельствуют высокие оценки наследуемости признаков, которые могут быть отобраны [148]. Тем не менее, имеется мало информации об адаптивном потенциале в средах, которым особенно угрожает изменение климата, таких как высокогорные районы.Одним из прогнозируемых последствий глобального изменения климата является перемещение видов растений на более высокие высоты и широты по мере смещения климата, к которому они приспособлены [149]. Резкое сокращение ареала или даже исчезновение видов растений может быть следствием миграционных процессов в сторону больших высот. Темпы и закономерности этой динамики будут сильно зависеть от предпочтений конкретного вида в среде обитания и от его функциональных особенностей [150]. Крутые экологические градиенты и неравномерное распределение местообитаний приводят к небольшому размеру и пространственной изоляции популяций и ограничивают поток генов [29].Было высказано предположение, что популяции видов растений могут сохраняться в своих нынешних ареалах и противостоять изменениям окружающей среды, если они обладают большой адаптивной способностью [151]. Гратани и др. [35], в сравнительном исследовании двух популяций Crepis pygmaea subsp. pygmaea и Isatis apennina , произрастающих на разных высотах в массиве Гран-Сассо (Италия), рассмотрели вопрос о том, может ли пластичность морфологических и физиологических признаков свидетельствовать об их будущем адаптивном потенциале для выживания при глобальном потеплении.Результаты подчеркивают, что C. pygmaea обладает значительно более высокой пластичностью как физиологических, так и морфологических признаков, чем I. apennina . Таким образом, предполагаемое повышение температуры воздуха может привести к тому, что C. pygmaea и I. apennina будут перемещены на более высокие высоты в массиве Гран-Сассо, при этом C. pygmaea будет благоприятствовать наибольшей пластичности. Исследования Sesleria nitida, , многолетнего травянистого вида, произрастающего на разных высотах вдоль узкого перепада высот в Центральных Апеннинах (Монте-Терминилло, Италия), показывают, что более низкие скорости фотосинтеза на большей высоте объясняются более сильным ветровым воздействием и более низкая влажность почвы на этом участке, а более низкий SLA (т.е., площадь наибольшей массы листа, LMA) способствует ограничению транспирации листа [65]. Степень, в которой SLA является фенотипически пластичным или генетически закрепленным, имеет важное значение для выживания популяций при изменении условий окружающей среды [152]. Кроме того, результаты показывают большую пластичность по физиологическим, чем по морфологическим и анатомическим признакам листа S. nitida , причем первый более полезен в условиях сильного стресса, изменяющегося в краткосрочной перспективе (табл. 3). Относительно большая фенотипическая пластичность S.nitida отражает его способность сохранять функцию в различных стрессовых условиях окружающей среды и выдерживать повышение температуры воздуха за счет потенциального смещения в сторону более высоких высот. Штёклин и др. [29] изучали последствия для альпийского ландшафта эволюционных процессов у четырех типичных альпийских видов растений ( Epilobium fleischeri , Geum reptans , Campanula thyrsoides и Poa alpina ). Внутрипопуляционное генетическое разнообразие четырех видов велико и в основном не связано с высотой и размером популяции.Тем не менее, генетическая дифференциация среди популяций сильно увеличивается с расстоянием, что предполагает значительный генетический дрейф среди популяций альпийских растений. Фенотипическая изменчивость формируется адаптивными, а также случайными эволюционными процессами, и пластические реакции на условия роста, по-видимому, имеют решающее значение для выживания растений в альпийском ландшафте.
PI PI PI PI Crepis Pygmaea Subsp. PYGMAEA La 0.2015 9061 0.65 09 LMA 0.38 0.53 Ltd 0.42 Wue 0.36 0,73 CHL 0.31RWC 0.21 0,55 Среднее значение 0,35 Среднее значение 0,50 91 529 Isatis apennina LA 0.25 DM 0.32 LMA 0.30 0.72 0.33 15 15 CHL 0,17 RWC 0.15 Среднее значение 5 0.30 среднее значение Sesteria Nitida Толщина листа 0.24 0,18 Высота главного бокового сосуда 0.30 9061 Ширина главных боковых сосудов 0,24 Высота главного бокового сосуда, 0.39 9061 0.32 0,47 Диаметр ксилематических сосудов 0,18 Mesophell плотность клеток 0,19 Толщина верхней склеренской слои 0.31 Общая площадь поверхности из чужевидных клеток 0.07 адаксиальная устичный длина 0,06 абаксиальной эпидермиса толщиной 0,06 адаксиальная Эпидермис толщиной 0,07 Leaf Mass Resear 0.15 Среднее значение 0.20 Среднее значение 64 Листовая площадь (LA), листовая сухая масса (дм), листовая площадь (LMA), плотность листьев (LTD ), чистый фотосинтез (), устьичная проводимость (), дыхание листа (), эффективность использования воды (WUE), водный потенциал листа в полдень (), общее содержание хлорофилла (Chl), относительное содержание воды (RWC), общий фотосинтез () , транспирация листьев (), соотношение между дыханием листьев и чистым фотосинтезом (). 7. Конкуренция между инвазивными и аборигенными видами
Повышение температуры воздуха и концентрации двуокиси углерода (CO 2 ) за последние десятилетия определило новые условия окружающей среды [153], которые могут действовать как мощный фактор естественного отбор среди видов, отдающий предпочтение более фенотипически пластичным видам [154] и приводящий к конкуренции между инвазивными и совместно встречающимися местными видами [153, 155]. Фенотипическая пластичность сосуществующих аборигенных и инвазионных видов в широколиственном лесу, формирующемся в природном заповеднике Сиро-Негри (45°12′39′′ с.ш., 9°3′26′′ в.д., Италия), свидетельствует о восприимчивости рассматриваемого вида к световым вариациям. . Q. robur (аборигенный вид) и R. pseudoacacia (инвазивный вид) обладают сходной физиологической пластичностью. Тем не менее, значительно более высокая морфологическая и анатомическая пластичность R. pseudoacacia , чем Q. robur , подтверждает его прошлую способность колонизировать лес и затем успешно врастать как в доминирующий, так и в доминирующий ярусы (данные Catoni et al. не опубликованы).
Интродуцированные виды часто проявляют небольшие генетические вариации в новой среде из-за генетического узкого места и дрейфа, характерного для небольших исходных популяций [156].Даже при генетических вариациях локальная адаптация может не возникнуть, если отбор слаб или непредсказуем, или если между популяциями происходит значительный поток генов [157]. Несмотря на эти ограничения, локальная адаптация часто способствует успеху растений, интродуцированных в новые условия [158–162]. В соответствии с этой гипотезой Niinemets et al. [42] демонстрируют более высокую пластичность инвазивного Rhododendron ponticum по сравнению с местным Ilex aquifolium в условиях высокой освещенности благодаря более высоким затратам азота на сбор света и фотосинтетический механизм, позволяющий производить большую площадь листа при одинаковом фотосинтезе и освещении. — характеристики урожая.Высокая фенотипическая пластичность в фотозащитных стратегиях и производительности инвазивного Taraxacum officinale повышает его конкурентоспособность в альпийской среде [68], учитывая, что интенсивность света является одним из наиболее меняющихся условий вдоль высотных градиентов. Растения T. officinale с больших высот, где условия освещения более изменчивы, обладают большей пластичностью, чем растения с более низких высот, что позволяет предположить, что пластичность экофизиологических признаков будет одной из основных стратегий колонизации сред с переменными условиями освещения.Высокая приспособляемость может быть связана либо с адаптивной стратегией, позволяющей справляться с колебаниями ресурсов в родном регионе [163, 164], либо с быстрой эволюцией в новых условиях после колонизации [165]. Дэвидсон и др. [153] показывают, что инвазивные виды более пластичны в своей реакции на большую доступность ресурсов, чем неинвазивные, но эта пластичность лишь иногда связана с преимуществом приспособленности, а неинвазивные виды сохраняют больший гомеостаз приспособленности при сравнении роста при низкой и средней доступности ресурсов.Функ [166] исследовал пластические реакции пяти инвазивно-местных пар в среде с низкими ресурсами на Гавайских островах и обнаружил, что максимальная скорость фотосинтеза и концентрация органического азота в листьях положительно связаны с приспособленностью инвазивных видов в ответ на доступность азота. Поскольку адаптивная пластичность может позволить определенным видам колонизировать экологически разнообразные районы без задержки, необходимой для местной адаптации, она может повысить их инвазивность и быстрое географическое распространение, способствуя вытеснению местных видов [4].
Тем не менее, несмотря на усилия последних десятилетий, эволюционные механизмы, ведущие к инвазивности, остаются неясными [167]. Молина-Черногория и др. [68] показывают, что инвазивные виды обладают значительно большей пластичностью, чем местные виды. Напротив, Дреновский и соавт. [168] предполагают, что местные и инвазивные виды могут конвергировать по функционально схожим признакам, демонстрируя сопоставимую способность реагировать на изменение доступности ресурсов. Кроме того, Годой и соавт. [169] показывают, что, несмотря на разумные аргументы в пользу связи фенотипической пластичности с инвазией растений, не было выявлено общего паттерна между фенотипической пластичностью и инвазивностью.ДеУолт и др. [170] проверили гипотезу о том, что тропический кустарник Clidemia hirta более распространен в интродуцированных (Гавайские острова), чем в естественном ареале (Коста-Рика) из-за генетических различий в приобретении ресурсов, распределении и фенотипической пластичности между местными и интродуцированными растениями. генотипы. Тем не менее результаты подчеркивают тот факт, что генетические сдвиги в использовании ресурсов, распределении ресурсов или пластичности не влияют на различия в распределении и изобилии среды обитания.Повышенная пластичность может не повышать приспособленность (неадаптивная пластичность) или даже снижать ее, а корреляция между различными признаками растений может придавать инвазивность за счет снижения стоимости неадаптивных и/или неадаптивных пластических признаков [14, 171]. Бастлова и Квет [43] показали фенотипическую изменчивость в местных популяциях Lythrum salicaria , евразийского вида, который успешно вторгся в водно-болотные угодья Северной Америки, конкурируя с местными видами растений [44]. Авторы показали, что растения, происходящие из более южных местонахождений Евразии, более похожи на инвазионные растения Северной Америки, чем на растения из северных местонахождений Евразии.Изменчивость характеристик роста по градиенту с севера на юг в пределах естественного ареала может быть результатом долговременной адаптации к преобладающим условиям окружающей среды, особенно к длине дня. Более того, изменчивость некоторых характеристик роста (например, сухой массы и количества боковых ветвей, сухой массы корней) как между евразийскими популяциями, так и внутри них указывает на пластическую реакцию роста на местные условия окружающей среды.
8. Выводы
В целом было показано, что изменение климата влияет на численность и распространение видов растений, а также на состав растительных сообществ [172, 173].Недавние исследования показывают, что при быстром изменении климата фенотипическая пластичность, а не генетическое разнообразие, вероятно, будет играть решающую роль в сохранении растений в окружающей среде [174]. Различная реакция на климат проявляется не только между популяциями в пределах ареала вида, но и между сосуществующими особями внутри популяции [149]. Тем не менее исследования генетической структуры на основе полиморфизма ферментов в популяциях лесных деревьев показали значительный уровень внутрипопуляционной изменчивости и небольшую межпопуляционную дифференциацию [61, 175, 176].Усиление летней засухи усугубит регенерацию многих видов деревьев в более низких широтных и высотных пределах их распространения, и считается, что интродукция более устойчивых к засухе видов в уязвимых местообитаниях способствует сохранению лесов [177]. Тем не менее интродукция более засухоустойчивых видов для смягчения последствий изменения климата не обязательно будет успешной из-за компромисса между засухоустойчивостью и пластичностью роста. Никотра и др. [178] подтверждают тот факт, что автохтонные происхождения обладают потенциалом устойчивости к изменению климатических условий в зависимости как от фенотипической пластичности, так и от генотипических вариаций.
Оценка уязвимости растительности и устойчивости к изменению климата требует понимания разнообразия видов растений в существующей растительности и стратегий их роста в ответ на колебания доступности ресурсов [179]. Поскольку виды с обширным географическим ареалом могут демонстрировать большие внутривидовые различия в физиологии, морфологии и фенологии, они могут быть хорошими моделями для изучения локальных и региональных адаптаций [140]. Тем не менее, адаптация к будущим глобальным изменениям может потребовать развития ряда различных признаков, которые могут быть ограничены корреляциями между ними [180].Таким образом, необходимо определить функциональные признаки растений, в которых пластичность, вероятно, будет определяющим фактором в реакции растений на глобальные изменения, способствуя прогнозированию изменений и сдвигов в распределении видов [178]. Кроме того, важно полностью понимать экологические последствия на уровне вида и экосистемы, учитывая, что виды с большей адаптивной пластичностью могут с большей вероятностью выжить в новых условиях окружающей среды, поскольку такие изменения обычно происходят слишком быстро, чтобы позволить эволюционный (или в в некоторых случаях миграционная) реакция.Пластичность признана основным источником фенотипических вариаций в реальном мире, поскольку она будет влиять на естественный отбор и, следовательно, на модели диверсификации среди популяций и, в конечном счете, видов [2, 181]. Пластичность способствует эволюционной диверсификации, если производимые фенотипы обеспечивают адаптивное разнообразие, которое при отборе становится эволюционно закрепленным [2, 182, 183]. Тем не менее, степень, в которой фенотипическая пластичность может способствовать выживанию в изменяющихся условиях окружающей среды, остается в значительной степени неизвестной.Хотя фенотипическая пластичность может способствовать кратковременной адаптации к изменениям окружающей среды, генетическая адаптация в конечном итоге может быть необходима для сохранения видов в экстремальных местообитаниях [177].
В ближайшем будущем будет важно собрать данные, работая в полевых условиях и, в частности, в девственных лесах и/или в хорошо сохранившихся местообитаниях, где новые факторы стресса ограничены, чтобы определить стандартные протоколы, полезные для сравнительных исследований. . Среди стратегий сохранение окружающей среды должно защищать неоднородность между средами обитания и внутри них, чтобы поддерживать большую внутривидовую изменчивость и тем самым сохранять множество фенотипических специализаций, которые смогут смягчить будущие экологические экстремальные явления из-за изменений климата и землепользования [184].
Конфликт интересов
Авторы заявляют об отсутствии конфликта интересов в отношении публикации данной статьи.
Происхождение механизма фенотипической пластичности глазных пятен сатироидной бабочки
[Примечание редактора: авторы повторно представили на рассмотрение исправленную версию документа. Далее следует ответ авторов на первый раунд обзора.]
Рецензент №2:
Мне очень понравилось читать эту рукопись Бхардваджа и его коллег.Это захватывающее, новое и поистине уникальное исследование. Сравнение пластических реакций с использованием реакций эво-дево у 12 видов беспрецедентно. Результаты показывают, что сложные гормональные взаимодействия, которые приводят к сезонной пластичности у бабочек Bicyclus, возникают поэтапно: чувствительность экдизона к температуре предшествует пластической реакции, в то время как экспрессия рецепторов экдизона в области глазного пятна и чувствительность к экдизону возникают в линия с сезонной пластичностью.Эти результаты имеют важное значение для нашего понимания эволюции пластичности и сложных черт в целом. Рисунки в этой рукописи являются прекрасной визуализацией основных результатов и закономерностей филогенетической изменчивости. Однако необходимы некоторые разъяснения в отношении трактовки пластичности и включения литературы, не относящейся к бабочкам.
Во-первых, рукопись в том виде, в каком она написана в настоящее время, сосредоточена главным образом на бабочках (например, начиная со второго абзаца), но результаты применимы к гораздо более широкому кругу систем.Было бы здорово, если бы соответствующие примеры из других систем можно было бы включить во Введение и Обсуждение. Например, работа Дж. У. Торнтона об эволюции рецепторов эстрогена касается как регуляторов пластичности (например, половых различий как пластичности развития), так и поэтапной эволюции сложных взаимодействий между сигнальными молекулами и рецепторами (например, преадаптация лигандов или рецепторов, см. и др., 2003 Наука, Торнтон, 2001 PNAS, Бриджем и др., 2006 Наука).
Спасибо.Теперь мы изменили введение, включив в него более общий абзац о происхождении пластичности в различных системах. Мы воздержались от включения приведенных выше ссылок (от Thornton et al.), потому что эти ссылки больше связаны с объяснением того, как семейство стероидных рецепторов достигло своего нынешнего разнообразия и как коэволюционируют системы рецепторных лигандов, что мы не исследуем в настоящей работе. текущая работа. Насколько нам известно, стероидный рецептор EcR присутствует у всех исследованных видов бабочек и в единственном экземпляре.
Во-вторых, рабочее определение пластичности в этой рукописи довольно специфично для полифенизмов и других форм эволюционных переключений (sensu MJ West-Eberhard). Это важно, потому что влияет на некоторые предположения во Введении и выводы в Обсуждении. Например, во Введении говорится, что «у нас все еще мало понимания того, как эти сложные адаптивные реакции возникают и развиваются в филогенезе», а в Обсуждении делается вывод, что «предупреждение о том, что если многие формы адаптивной пластичности столь же специфичны и трудно эволюционируют, как один задокументирован в Б.anynana, эти изысканные приспособления к конкретным предсказуемым флуктуирующим условиям окружающей среды могут на самом деле сделать вид уязвимым к вымиранию». эти утверждения кажутся менее применимыми к другим формам пластичности. Например, Ньюман и Мюллер утверждали, что пластичность — с точки зрения чувствительности к окружающей среде — является наследственным состоянием для большинства фенотипов, и что канализация развития изначально индуцированных окружающей средой планов тела происходит (2000, «Эпигенетические механизмы возникновения характера»).В другом случае рассмотрим обучение методом проб и ошибок как тип пластичности — существует множество исследований, посвященных размеру мозга и обучению в разных филогенезах, и эти формы пластичности значительно менее «хрупки» по отношению к производительности в полной мере. новые условия. Я предполагаю, что рукопись построена конкретно вокруг полифенизмов или эволюционных молекулярных взаимодействий, связанных с пластичностью.
Да, мы согласны с тем, что существуют разные взгляды на происхождение пластичности и на то, является ли пластичность производным или исходным состоянием для большинства организмов.По нашему мнению, обе точки зрения на происхождение пластичности на самом деле возможны, но важно в каждом случае использовать филогенетический сравнительный метод для изучения наследственных состояний и направления изменений, что редко делается, особенно при изучении эволюции пластических систем. на молекулярном, ближайшем уровне. Мы включили абзац ниже, чтобы представить различные взгляды, которые преобладают в области пластичности, а затем объяснить, что наше внимание будет сосредоточено на изучении происхождения адаптивной пластичности, лежащей в основе полифенизма рисунка крыльев у сатирид, представленных Bicyclus anynana :
.«Существуют два разных взгляда на фенотипическую пластичность.Один рассматривает пластичность как производную адаптацию, помогающую организмам выживать в изменяющихся условиях (Bradshaw, 1965; de Jong, 2005), в то время как другой рассматривает пластичность как результат гибких, неканализованных процессов развития, изначально присутствующих у большинства организмов, которые помогают они колонизируют или адаптируются к новой среде, например, преадаптация (Newman and Müller, 2000; West-Eberhard, 2003; Pigliucci, 2005; Laland et al., 2014). […] Здесь мы сосредоточим наше исследование на механистическом происхождении адаптивного сезонного полифенизма, когда сигналы окружающей среды, испытываемые во время развития, изменяют фенотипы взрослых, чтобы они соответствовали различным сезонным повторяющимся условиям, что является высокоразвитой формой фенотипической пластичности.
Что касается методов, у меня есть только одна незначительная проблема. Я понимаю трудности выращивания 12 видов бабочек, особенно когда им требуются разные растения-хозяева и условия выращивания. Однако этот вид выращивался в трех разных местах, а Bicyclus anynana выращивался в уникальном месте. Было бы целесообразно обсудить в разделе «Материалы и методы» возможную роль изменчивости в среде выращивания – есть ли шанс, что различия между видами могут быть связаны скорее с окружающей средой, чем с характеристиками вида?
Спасибо.В наших методах мы объяснили, что эти бабочки выращивались в камерах с контролируемым климатом во всех трех местах, где мы следили за температурой и влажностью во время развития личинок. Факторы окружающей среды были одинаковыми в разных местах, что означает, что их влияние в этом исследовании минимально.
Рецензент №3:
В этой рукописи проводится сравнительный анализ контролируемой температурой пластичности размеров пятен на крыльях бабочки («глазных пятен»).В исследовании делается попытка применить филогенетический подход, чтобы сделать вывод об эволюционной истории различных аспектов механизма температурной реакции глазного пятна. Авторы рассматривают влияние температуры выращивания на размер глазного пятна у 13 видов, что является интересным и ценным упражнением. Они также изучают влияние температуры на титры гормона (20E), влияющего на полифенизм окраски у некоторых видов бабочек, а также на экспрессию EcR, предполагаемого рецептора 20E. Наконец, они рассматривают эффекты инъекции 20E, а также антагониста рецептора в один момент времени у четырех видов.На основании этих сравнительных наборов данных они делают филогенетический аргумент в пользу того, что пластичность глазных пятен у одного конкретного вида произошла в результате постепенной сборки нового механизма гормональной реакции. Мои общие мысли таковы, что мотивация работы очень интересна, и здесь есть некоторые ценные данные. Но в исследовании также есть многочисленные фатальные недостатки с точки зрения дизайна эксперимента и качества данных (описанных ниже), которые заставляют меня весьма скептически относиться к интерпретации данных авторами.
Важные выводы статьи основаны в основном на отрицательных результатах. Моя самая сильная критика всего этого исследования, как и многих других работ evo-devo, посвященных анализу филогенетических признаков, заключается в том, что основные выводы основаны на плохо контролируемых отрицательных данных.
Спасибо за ваш комментарий. Отрицательные результаты (подкрепленные соответствующими контролями) столь же информативны, как и положительные результаты. Рецензент предположил, что данные были построены на плохо контролируемых отрицательных данных, но это не так.Мы включили дополнительную информацию о том, как мы контролировали эти отрицательные результаты, в разделе «Материалы и методы» рукописи.
На самом деле почти для каждого признака, который подлежит филогенетическому анализу в этой статье, у меня есть серьезные опасения по поводу проблем с плохой выборкой временных рядов, выборкой на неправильной стадии развития, проведением гомологии между видами и т. д. И беспокойство со всеми эти вопросы являются отрицательными данными. Отрицательные результаты от разных видов учитываются как состояния признаков в филогенетическом анализе, что приводит авторов к выводу об эволюции механизма развития.Но, как я описываю ниже, есть веские основания полагать, что отсутствие данных временных рядов (и в одном случае явно преднамеренное игнорирование опубликованных данных) смещает эту работу в сторону получения отрицательных результатов.
Мы считаем эту критику несправедливой и ошибочной. Также неясно, что рецензент считает «отрицательными результатами». Мы не преследовали какой-либо конкретный результат при подготовке к проведению этого исследования. Мы просто сообщаем о том, что нашли. Кроме того, эта работа представляет собой наиболее подробное исследование чешуекрылых, проведенное на сегодняшний день и включающее тщательно отобранные образцы на гомологичных стадиях развития.Первоначально мы использовали покадровую фотографию, чтобы установить стадию Wr как чувствительную к температуре стадию пластичности размера вентрального пятна Cu1 в модельной системе ( Bicyclus anynana ) (Monteiro et al., 2015). Личинки помещались по отдельности в прозрачные вертикальные емкости с кормом и фотографировались каждые 15 минут. Когда личинки покидали корм и поднимались по контейнеру вверх (не возвращаясь), они маркировались как достигшие начала стадии блуждания. Мы выполнили такую же покадровую съемку для всех других видов.Количественная оценка титров 20E, окрашивания EcR и инъекций 20E выполнялась на основе этой тщательной стадийной стадии блуждания, чтобы позволить нам сравнить действительно гомологичные точки развития у всех видов. Большая часть этой информации уже была подробно описана в методах в исходной заявке, но мы сделали на этом еще больший акцент.
Известно, что фотопериод влияет на пластичность рисунка крыльев, но не контролировался в зависимости от обработки и вида. Фотопериод выращивания у некоторых видов различен, а для многих других видов просто не сообщается (я не могу найти информацию в рукописи).Это серьезно искажает результаты, представленные на рис. 1, поскольку известно, что фотопериод оказывает сильное влияние на черты пластического рисунка крыльев у многих видов бабочек, как независимо от температуры, так и во взаимодействии с ней. Без контроля фотопериода невозможно определить, какая часть фенотипической реакции, показанной на рисунке 1, обусловлена температурой по сравнению с другими факторами окружающей среды. Чтобы быть опубликованным, для всех видов фотопериод должен быть одинаковым для обеих температурных обработок, и фотопериод должен быть указан в статье.
Спасибо, что указали на это важное упущение. Теперь мы изменили сведения о материалах и методах, чтобы включить эту информацию:
«Все виды выращивались при двух температурах, разделенных на 10 градусов, при относительной влажности 70 или 80% и при цикле свет:темнота 12:12. Единственным исключением была Junonia coenia, , которую выращивали при цикле свет:темнота 16:8. […] Влажность в этих последних камерах контролировали с помощью гигрометра PT2470 (Exoreptiles, Малайзия) и регистраторов данных EL-USB-2 (Lascar Electronics, PA 16505, США).
Нет никаких доказательств того, что «глазные пятна» Papilio, Danaus и Idea имеют общую эволюционную или эволюционную гомологию с настоящими нимфалидными глазными пятнами. Мне неизвестны какие-либо данные о развитии, экспрессии генов или генетические данные, подтверждающие гомологию этих цветовых узоров с настоящими пятнами на глазах. По моему мнению, эти паттерны, скорее всего, происходят от центральной или маргинальной систем формирования паттернов WntA. Они выглядят точно так же, как паттерны, утраченные при нокауте WntA CRISPR у других видов (Mazo-Vargas et al., 2017). Если указание авторов на гомологию неверно, то любые данные, полученные в результате анализа этих цветовых паттернов, полностью исказят филогенетический анализ. Эта потенциальная проблема особенно критична, поскольку это внешние группы.
Предыдущая работа предполагала, что глазные пятна произошли от существующих пятен, возможно, подобных тем, что показаны на Idea , Daninae (подсемейство Nymphalidae с базальным разветвлением) (Oliver et al., 2014). Из-за этой предыдущей работы мы решили также изучить виды с более простыми пятнами внутри и вне Nymphalidae.Ген Spalt является известным маркером пятен и пятен на глазах у Nymphalidae и Pieridae (см. рисунок S2 (Oliver et al., 2012) и (Stoehr et al., 2013)) и теперь мы изменили рисунок 2, включив в него Spalt co. — окрашивание, выполненное одновременно с EcR для всех видов. Мы наблюдаем, что пятна, локализованные в гомологичных позициях как глазки Cu1, у Papilio и Idea экспрессируют Spalt во время развития точно так же, как и глазные пятна. Этот общий паттерн экспрессии предполагает (но не доказывает окончательно), что глазные пятна и пятна могут быть гомологичны, и что пятна могли предшествовать глазным пятнам в эволюции.Важно, однако, то, что все виды с пятнами внешней группы не экспрессировали EcR в пятнах или в какой-либо области, которая могла бы быть гомологичной пятнам на глазах, мы пришли к выводу, что паттерн экспрессии EcR, вероятно, возник в сестринской линии по отношению к Daninae, т.е. также линия происхождения глазных пятен. У Danaus , другой Danainae, пятна на краях крыльев действительно, вероятно, представляют собой систему формирования паттернов на краях крыльев, на которую влияет WntA, который может не быть гомологичным ни пятнам (как в Idea ), ни пятнам-глазам.Несмотря на это, этот результат, вопреки тому, что предполагает рецензент, не «полностью искажает филогенетический анализ». Включение этого вида не оказывает существенного влияния на нашу реконструкцию наследственного состояния происхождения экспрессии EcR в областях, которые картируются в центрах глазных пятен.
Пятна антител
EcR имеют низкое качество, не имеют надлежащего контроля и предназначены только для одного этапа. Иммунные пятна EcR на рисунке 2 являются важными данными для построения филогенетической модели. К сожалению, с этими данными много проблем, и они не могут быть опубликованы, как показано.Прежде всего, нет положительных контролей для видов, которые оцениваются как не имеющие иммунофлуоресцентного сигнала. Знаем ли мы, что антитела работают у этих видов?
Теперь мы включили новую дополнительную фигуру (рис. 2 — дополнение к рисунку 2), чтобы включить положительные контроли экспрессии EcR в больших ядрах периподиальной мембраны, как сообщалось ранее для EcR (Koch et al., 2003). Кроме того, мы добавили в основной рис. 2 совместное окрашивание Spalt (Sal), потому что этот ген маркирует будущие центры пятен и глазных пятен.Экспрессия Sal присутствовала у всех видов, исследованных с пятнами/глазками, независимо от того, экспрессировали ли они EcR в этих элементах паттерна.
Во-вторых, временные ряды выражений отсутствуют. Возможно, в отрицательных случаях экспрессия EcR немного раньше или позже. Я очень озадачен этим отсутствием учета других моментов времени, кроме стадии блуждания личинки.
В предыдущей работе мы установили, что стадия блуждания является критически чувствительной стадией развития, когда размер вентрального глазного пятна Cu1 определяется у B.anynana в ответ на температуру (Monteiro et al., 2015). Это было отправной точкой нашего сравнительного исследования. За всеми животными, использованными в наших экспериментах, наблюдали на протяжении всего их развития, и мы смогли с уверенностью идентифицировать одну и ту же гомологичную стадию блуждания, используя замедленную съемку у всех видов. Расширение текущей работы (с 13 видами) для включения выборки окрашивания EcR в другие моменты времени было бы непомерно с точки зрения усилий по выборке и не требуется для ответа на вопрос, который мы поставили в начале этой рукописи.Наша работа не исключает будущих исследований этих альтернативных моментов времени как возможных стадий развития, которые регулируют пластичность у других видов. Обратите внимание, однако, что часто разные цис-регуляторные элементы контролируют экспрессию одного и того же гена в разных местах тела и даже на стадиях развития в гомологичных местах. Известным примером последнего являются различные цис-элементы гена Distal-less , которые инициируют экспрессию в ранних конечностях Drosophila , а затем поддерживают эту экспрессию на более поздних стадиях развития конечностей (McKay et al., 2009). Это подразумевает, что происхождение новых цис-элементов или разрушение гомологичных и ранее существовавших цис-элементов может быть ответственным за изменение паттерна экспрессии генов в ходе развития в гомологичных положениях крыла в течение эволюционного времени. Чтобы избежать этих смешанных эффектов, мы ограничили наше исследование молекулярной эволюции механизмов пластичности у Bicyclus anynana , взяв образцы строго гомологичных тканей (вентральные крылья и секторы крыльев Cu1) и временные точки в филогенезе.
Например, экспрессия EcR была опубликована в глазных пятнах Junonia (Precis) coenia в более поздний момент времени, в не меньшей степени коррелируя с фактором транскрипции глазных пятен Distalless (Koch et al., 2003). Но авторы полностью игнорируют это в своей модели и даже оценивают экспрессию EcR J. coenia как отрицательную в своем филогенетическом анализе. Я думаю, что это игнорирование различных моментов времени и ранее опубликованных данных удивительно и, надеюсь, не вводит в заблуждение намеренно.
Рецензент ошибся, полагая, что мы проигнорировали этот предыдущий результат.Нам хорошо известно об этом предыдущем исследовании, но по причинам, уже изложенным выше, мы решили сосредоточить наше исследование на строго гомологичных периодах развития у всех исследованных видов. Относительно J. coenia , предыдущая работа задокументировала экспрессию EcR в периподиальной мембране и вдоль трахеи, но не в фокальных центральных клетках глазного пятна на блуждающей стадии развития (Koch et al., 2003). EcR позже наблюдается в глазных пятнах J. coenia во время развития куколки, что является более поздней стадией и не является предметом этого исследования.Учитывая, что близкие родственники, такие как J. atlites и J. iphita , имеют экспрессию EcR в вентральных центрах глазных пятен Cu1 на блуждающих стадиях развития, наиболее вероятное эволюционное объяснение наблюдаемого отсутствия EcR в глазных пятнах у этого вида и также у J. almana наблюдается потеря этого паттерна экспрессии на этой стадии развития. Обратите внимание, что этот тип специфической для стадии регуляции EcR был документирован ранее у B. anynana вентральных глазных пятен на переднем крыле (только у самок; Monteiro et al., 2015), где EcR экспрессируется на средних личиночных стадиях и снова на ранних стадиях куколки, но отсутствует на стадии блуждания, что позволяет этим центрам глазных пятен быть нечувствительными к температуре. Это позволяет этим глазным пятнам сохранять одинаковый размер как в сезонных формах, так и круглый год участвовать в половом отборе и / или избегании хищников. Опять же, мы не игнорируем другие временные точки (они просто выходят за рамки текущего исследования) и не намеренно вводим читателей в заблуждение.
Я также нахожу очень странным, что нет J.coenia EcR иммуноокрашивает в этой статье, тем более, что один из экспертов по видам (Х. Нейхаут) является соавтором статьи. Как я неоднократно указывал в этом обзоре, авторам необходимо учитывать более чем одну стадию разработки, чтобы адекватно ответить на вопросы своего исследования.
Хотя рецензент прав, указывая, что H. Nijhout является экспертом и соавтором текущего исследования, мы тщательно изучили существующую литературу (выполненную Koch и Nijhout) и использовали их конкретные данные экспрессии EcR из J.coenia (Koch et al., 2003), чтобы провести реконструкцию состояния наших предков.
В-третьих, некоторые пятна выглядят некачественно. Мне они не кажутся точечной экспрессией, а размер изображения, кадрирование и разрешение плохие, поэтому я не могу сказать, что сигнал локализован в ядре, как можно было бы ожидать для ЭКР. Я хотел бы увидеть дополнительный рисунок с изображениями всего крыла для всех видов, чтобы более четко увидеть корреляции цветового рисунка и исключить фоновую флуоресценцию.Я хотел бы видеть правильное контрастное окрашивание. Я хотел бы видеть положительный контроль антител EcR у каждого вида. Я хотел бы увидеть изображения с большим увеличением, показывающие предполагаемые «точечные» узоры, а также ядерные локализации. Если возможно, это также сделало бы данные намного более убедительными, чтобы увидеть двойные пятна для проверки совместной локализации EcR и некоторых других маркеров глазных пятен. Это было бы особенно важно для некоторых видов, у которых пятна больше похожи на размытые пятна на предоставленных изображениях.
Теперь мы включили двойное окрашивание для гена положительного контроля, Spalt, ранее показанного для маркировки пятен и глазных пятен, а также вспомогательное изображение для рисунка 2, которое включает положительные контроли для окрашивания EcR в больших ядрах периподиальной мембраны для всех виды, за исключением одного (у которого в то время не было сфотографировано периподиальной мембраны и образцы которого с тех пор испортились). Однако этот вид имел положительный сигнал EcR в центрах глазных пятен. Клетки крылового диска личинки очень маленькие, и точечное окрашивание ядер очень трудно визуализировать.Однако окрашивание ядер в полиплоидных ядрах периподиальной мембраны легко визуализировать и сделать соответствующие положительные контроли для окрашивания EcR. Кадрирование и разрешение представленных изображений (включая полное поле зрения, полученное с помощью 40-кратного объектива для всех образцов) были необходимы для надлежащего конфокального среза только вентральной поверхности крыла, предмета текущего исследования. Более низкое разрешение затруднило бы выделение только вентрального сигнала от любого сигнала, присутствующего на дорсальной поверхности крыла, которая не исследовалась.Обратите внимание, что для получения такого типа данных, специфичных для поверхности, мы проводили рассечение левого и правого крыла отдельно, чтобы всегда иметь возможность различать дорсальную и вентральную поверхности крыльев для каждого исследованного экземпляра.
Одноэтапный отбор проб для сравнения титра гормонов не имеет смысла, поскольку известно, что критические периоды различаются у разных видов. Титры 20E в этом исследовании были предоставлены только для одной временной точки (бродячие личинки), потому что это важная стадия для B. anynana.Есть несколько важных проблем с этим отсутствием временной выборки. Во-первых, у насекомых синхронизация импульсов гормонов и периодов чувствительности может быть очень специфичной и недолговечной, и небольшие изменения в этих импульсах и периодах почти наверняка имеют значение для эволюции признаков. Поэтому один момент времени, как правило, не может служить показателем всего эндокринного процесса в межвидовом сравнительном исследовании, по крайней мере, без какого-либо эталонного временного ряда для каждого вида. Во-вторых, это почти наверняка является проблемой в данном исследовании, поскольку известно, что импульс 20E и чувствительный период для полифенизма цветовых паттернов у J.coenia относится к более позднему периоду времени и даже не рассматривается в этой статье (Rountree and Nijhout, 1995). Я обеспокоен тем, что при отборе проб только в один момент времени у всех видов в этом исследовании могут быть важные ложноотрицательные результаты. В-третьих, я думаю, что, не делая временных рядов, просто упускается много интересной биологии. Для меня очень интересно, что и B. anynana, и J. coenia экспрессируют EcR в своих глазных пятнах (последнее наблюдение в основном игнорируется в этой статье), но имеют разные чувствительные периоды и реакции.Ясно, что здесь происходит более сложная эволюция. Я хотел бы, чтобы в этой статье мы больше говорили об этом. Самое главное, однако, чтобы у них был титровый временной ряд, чтобы получилась полная, честная и интересная история.
Да, мы согласны с автором обзора в том, что эволюция критического периода времени для чувствительности к температуре у разных видов является вероятным объяснением того, почему другие виды не проявляют особой формы пластичности, встречающейся у B. anynana . Однако наша цель состояла в том, чтобы попытаться изучить, как конкретный образец пластичности в B.Anynana эволюционировала. Мы не собирались объяснять, как эволюционировали все паттерны пластичности, которые могли присутствовать у всех этих видов. Это было бы громоздкой целью для одного исследования. Кроме того, мы не можем проводить дополнительную временную выборку на данном этапе из-за экспериментальных и финансовых ограничений. Мы изменили нашу рукопись в определенных разделах, чтобы сделать наши цели более ясными, а также отметить, что критический период времени для чувствительности к температуре, вероятно, меняется у разных видов.
Постановка всех экспериментов по инъекциям на одном этапе недостаточна для сравнительного анализа, поскольку известно, что периоды гормональной чувствительности у разных видов различаются. Все инъекции 20E и CucB были на одной стадии (блуждающая личинка). Это проблематично, потому что многие ключевые выводы статьи основаны на отрицательных результатах этих инъекций. По многим причинам, изложенным выше, включая предыдущие экспериментальные данные J. coenia, очень важно делать несколько моментов времени.Исходя из этих инъекций в один момент времени, мы не можем исключить возможность того, что невосприимчивые виды могут просто реагировать в разные моменты времени. Я особенно скептически отношусь к V. dejone, у которого наблюдается экспрессия EcR в глазном пятне и очень сильный температурный отклик глазного пятна. Как я описывал в другом месте, я также скептически отношусь к тому, что черные пятна у I. leucone вообще гомологичны пятнам на глазах B. anyana, поэтому я не уверен, насколько значимы результаты для этого вида.
Да, мы согласны с рецензентом в том, что мы не можем исключить возможность того, что другие виды регулируют свои паттерны пластичности на разных стадиях развития, но это не то, что мы собираемся исследовать.Опять же, мы сосредоточили наше исследование на стадии блуждания, где существуют определенные температурно-опосредованные различия в титрах гормонов и передаче сигналов EcR у Bicyclus anynana , видов, вокруг которых мы сосредоточили это исследование.
Странное несоответствие в управлении впрыском. Не должны ли отрицательные контроли «V» быть одинаковыми между экспериментами 20E и CucB? Тем не менее, они очень разные, в некоторых случаях требующие совершенно разных масштабов осей между экспериментами.В некоторых случаях различия между различными отрицательными контролями намного больше, чем различия между лечением и отрицательным контролем. Результаты B. anyana выглядят особенно странно. Либо я упускаю что-то, что нужно объяснить получше, либо были какие-то проблемы с экспериментами.
В то время как отрицательным контролям «носитель» действительно вводили один и тот же этанол-солевой раствор, мы выполнили две серии инъекций для животных, которым вводили носитель.Один выполнялся вместе с инъекциями 20E, а другой — вместе с инъекциями CucB. Эти две группы транспортных средств были разделены для согласованности. Небольшие различия в «V» связаны с разными партиями личинок, использованных для экспериментов.
https://doi.org/10.7554/eLife.49544.sa2Что такое пластичность нейронов и почему это важно?
Нейрональная пластичность или нейропластичность — это биологический процесс, посредством которого мозг реорганизует свои синапсы в ответ на раздражители.Мозг всегда стремится оптимизировать свое функционирование, и поэтому существуют биологические системы, которые часто усиливают задействованные модели мышления, делая соответствующие нейронные сети сильнее, а ненужные — ослабляя, поддерживая такие процессы, как обучение и память.
Изображение предоставлено: Андрей Водолажский / Shutterstock.com
Исследование основных биологических механизмов нейропластичности оказалось неоценимым, поскольку помогло ученым понять не только то, как обучение и память происходят в мозге, но также предоставило модель некоторых нервно-психических расстройств и важные сведения о восстановлении после черепно-мозговой травмы.Из-за своего значения в этих ключевых областях психического здоровья нейропластичность считается важной областью изучения.
Что такое пластичность нейронов?
Нейропластичность относится к биологическому процессу, при котором активность мозга и реакции на стимулы влияют на силу или слабость конкретных синаптических связей между синапсами или даже создают новые связи.
Кульминацией этой синаптической пластичности является нейропластичность, поскольку усиление релевантных и ослабление нерелевантных связей позволяет мозгу динамично реагировать на множество ситуаций и помогает ему стать эффективным и реагировать на повторяющиеся стимулы.
Эти изменения в определенных синаптических связях способствуют изменениям в более крупных нейронных сетях, которые могут отвечать за различные функции. Например, эти нейронные сети могут определять, как мы ведем себя в определенных сценариях, и они могут формироваться в результате обучения или помогать нам создавать воспоминания. Пластичность нейронов также связана с различными неврологическими расстройствами, и появляется все больше свидетельств того, что дисфункция этого механизма может привести к различным патологиям.
Значение нейропластичности
Нейропластичность невероятно важна, потому что она лежит в основе основных человеческих переживаний. Обучение и память, ключевые составляющие того, что значит быть человеком, были бы невозможны без этого процесса. Кроме того, продолжение изучения нейропластичности считается необходимым для расширения наших знаний о нервно-психических расстройствах и травмах головного мозга и поможет найти новые и улучшенные варианты лечения.
Значение пластичности нейронов хорошо иллюстрируется исследованиями, проведенными с участием буддийских монахов.Чуть более десяти лет назад ученые работали с Далай-ламой, который помог им нанять тибетских буддийских монахов для многочисленных исследований того, как медитация влияет на физический мозг. На протяжении веков эта практика была хорошо известна своей способностью воздействовать на разум, но до этого момента было мало свидетельств того, как медитация может изменить нейронные схемы мозга.
Многочисленные исследования были проведены в Лаборатории визуализации мозга и поведения Вайсмана в Университете Висконсин-Мэдисон, и результаты оказались плодотворными.Ученым удалось доказать, что медитация может вызывать физические изменения в мозге, изменяя его структуру и функции. Результаты можно использовать в качестве признака потенциала нейропластичности, поскольку исследование показывает, насколько податливым является мозг и насколько он восприимчив к изменениям только благодаря умственным усилиям.
Ниже описывается роль нейропластичности и значение ее изучения в различных видных областях психологических исследований.
Нейропластичность и депрессия
Нарушения структурной пластичности связаны с развитием большой депрессии.Исследование показало, что эффективность антидепрессантов может быть, по крайней мере частично, связана с их влиянием на коррекцию нарушений структурной пластичности.
Действительно, исследования показали, что ингибиторы обратного захвата серотонина (СИОЗС) играют роль в опосредовании нейропластичности, подобной развитию. Эти результаты означают, что будущие методы лечения могут быть разработаны с учетом этой модели нарушения пластичности.
Нейропластичность и обучение
Растущее число исследований подтверждает роль нейропластичности в обучении.Способность мозга к перенастройке делает возможным приобретение новой информации и новых навыков. Эта новая информация должна воздействовать на мозг, чтобы усвоить ее: природа обучения заключается в том, что мы меняемся благодаря опыту.
Нейровизуализационные исследования подтвердили, что анатомические изменения происходят в мозгу тех, кто изучает второй язык. Эти результаты показывают нейропластичность в действии, изменяя структуру мозга, чтобы обеспечить возможность обучения.
Нейропластичность и инсульт
Исследования показали, что мозг полностью способен динамически реагировать на ситуации, которые позволяют ему вносить изменения в свои нейронные цепи.Это необходимо для восстановления после черепно-мозговой травмы, например, инсульта. Исследования показали, что нейропластичность частично отвечает за успешное восстановление после инсульта.
Имеющиеся данные убедительно свидетельствуют о том, что наилучшие результаты восстановления обеспечивают реабилитационные тренировки, которые повышают пластичность нейронов, помогая мозгу пациентов восстановить функции за счет изменения его схемы. Расширяя наше понимание этого, ученые могут разрабатывать новые методы лечения, которые могут помочь улучшить результаты лечения.
Нейропластичность и когнитивно-поведенческая терапия
Было показано, что когнитивно-поведенческая терапия (КПТ) не менее эффективна в лечении депрессии, чем лекарства. Он также считается очень эффективным при лечении тревожных расстройств.
Недавние исследования показали, что когнитивно-поведенческая терапия оказывает влияние на физические изменения мозга, подчеркивая роль нейропластичности в успешном выздоровлении от этих психологических расстройств. Например, одно исследование показало, что у людей с тревожными расстройствами наблюдается повышенная активность миндалевидного тела.После когнитивно-поведенческой терапии объем серого вещества в этой области уменьшился, как и нейронная чувствительность. Полученные данные подтвердили, что успех когнитивно-поведенческой терапии связан со структурными и активационными изменениями, вызванными пластичностью нейронов.
Резюме
Биологический процесс пластичности нейронов позволяет изменять нейронные схемы, что может изменить структуру и функции мозга. Эти изменения позволяют обучению и памяти работать правильно, а отклонения в ее работе связаны с нервно-психическими расстройствами.Изучение пластичности нейронов важно из-за ее значения для лечения этих расстройств, а также для лечения черепно-мозговой травмы.
Источники:
- Дэвидсон Р. и Лутц А. (2008). Мозг Будды: нейропластичность и медитация [в центре внимания]. Журнал обработки сигналов IEEE, 25 (1), стр. 176–174. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2944261/
- Фукс, Э., Чех, Б., Коле, М., Михаэлис, Т., и Лукассен, П. (2004).Изменения нейропластичности при депрессии: гиппокамп и не только. Европейская нейропсихофармакология, 14, стр. S481-S490. www.sciencedirect.com/science/article/abs/pii/S0924977X04001269
- Ли, П., Лего, Дж., и Литкофски, К. (2014). Нейропластичность как функция изучения второго языка: анатомические изменения в человеческом мозгу. Кортекс, 58, стр. 301-324. https://www.sciencedirect.com/science/article/pii/S0010945214001543
- Монссон, К., Салами, А., Фрик, А., Карлбринг П., Андерссон Г., Фурмарк Т. и Бораксбекк К. (2016). Нейропластичность в ответ на когнитивно-поведенческую терапию социального тревожного расстройства. Трансляционная психиатрия, 6 (2), стр. e727-e727. https://www.nature.com/articles/tp2015218
- Восс, П., Томас, М., Сиснерос-Франко, Дж., и де Виллер-Сидани, Э. (2017). Динамический мозг и меняющиеся правила нейропластичности: значение для обучения и восстановления. Frontiers in Psychology, 8. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5649212 .
Дополнительная литература
Что такое нейропластичность? — Определение и концепция — Видео и стенограмма урока
Как работает нейропластичность?
Процесс нейропластичности не быстрый и не простой; скорее, это происходит на протяжении всей вашей жизни и может включать множество процессов.Наряду с изменением ваших нейронных синапсов и проводящих путей это может включать изменения в ваших нейронах, сосудистых клетках и глиальных клетках. Нейропластичность также происходит рука об руку с синаптической обрезкой, которая представляет собой способ мозга удалить нейронные связи, которые больше не нужны или полезны, и укрепить необходимые. То, как ваш мозг решает, какие связи обрезать, зависит от вашего жизненного опыта и от того, как давно вы использовали связи. Точно так же нейроны, которые становятся слабыми из-за недостаточного использования, отмирают в процессе апоптоза .В общем, нейропластичность — это способ настроить ваш мозг на эффективность.
Нейропластичность происходит постоянно по мере того, как вы изучаете и запоминаете новые данные, а также по мере развития вашего мозга; однако это также может быть вызвано физической травмой. В таких случаях нейропластичность служит адаптивным механизмом, позволяющим компенсировать потерю функций после телесных повреждений. Например, если кто-то получает черепно-мозговую травму, нейропластичность позволяет мозгу «перемонтировать» себя, чтобы восстановить или максимизировать функционирование мозга, перестраивая нейронные цепи и позволяя неповрежденной части мозга взять на себя поврежденную часть.
Роль нейропластичности в депрессии
Давно известно, что расстройства настроения связаны с нейрохимическими изменениями; тем не менее, связь между нейропластичностью и депрессией до недавнего времени оставалась неизведанной территорией. Исследователи обнаружили, что депрессия, хронический стресс (который может усугубить депрессию) и другие расстройства настроения могут снижать нейропластичность, что приводит к уменьшению количества синаптических связей.Это, в свою очередь, может привести к снижению шансов на выживание клеток, а также к снижению эффективности нервных синапсов. И наоборот, было обнаружено, что лечение антидепрессантами создает противоположный эффект — повышает нейропластичность.
Конечно, эти результаты опровергают устаревшее мнение о том, что каждый человек рождается с определенным количеством клеток мозга и что, как только клетка умирает, она исчезает навсегда. Вместо этого мозг продолжает изменять свои синапсы и проводящие пути и выращивать новые клетки. Даже во взрослой жизни ваш гиппокамп и другие области мозга могут продолжать выращивать новые клетки мозга.
Однако нейропластичность человека зависит от его или ее повседневной деятельности. Например, британский исследователь Элеонора Магуайр изучала лондонских таксистов, которым приходилось запоминать сложную карту Лондона, чтобы получить права. Она обнаружила, что после завершения интенсивного и продолжительного исследования, связанного с заучиванием карты, у тех, кто дольше всех водил машину, наблюдалось наибольшее улучшение в области гиппокампа мозга. Также было высказано предположение, что здоровый режим упражнений может положительно повлиять на нейропластичность.
Краткий обзор урока
Нейропластичность — это изменение нервных путей и синапсов, происходящее из-за определенных факторов, таких как поведение, окружающая среда или нервные процессы. Во время таких изменений мозг занимается синаптической обрезкой , удаляя ненужные или бесполезные нейронные связи и укрепляя нужные.